Clear Sky Science · zh
旋转细菌鞭毛产生的流体力矩偶极驱动对称圆盘
能带动微小齿轮旋转的小游动体
乍一看,一团游动的细菌像是漫无目的的混乱。研究表明,在特定条件下,这些微生物却能做出令人意外的有序行为:它们可以让完全圆形、对称的圆盘朝任意选定方向旋转,而无需直接推动圆盘的边缘。该工作展示了如何收集细菌鞭毛的扭转运动作为一种新型的微观动力来源,对智能材料、微型机器以及细菌在狭小空间中运动的理解都可能产生影响。
细菌如何运动并搅动周围流体
像大肠杆菌(Escherichia coli)这样的有运动能力的细菌通过旋转马达来驱动长而柔软的尾状结构——鞭毛。在几微米的尺度下,水的行为更像浓稠的糖浆:没有滑行,细胞要保持移动就必须不断对流体施力。多年来,物理学家主要用细菌沿运动方向拉动或推动流体的观点来描述游动微生物,这一图景能解释邻近颗粒扩散增强,甚至稠密微生物溶液表现出类似“超流体”的现象。但这种标准描述大多忽略了运动的另一个特征:因为鞭毛束朝一个方向旋转而细胞体为平衡总力矩朝相反方向旋转,每个细菌实际上还作用像一对反向旋转的微小搅拌器。
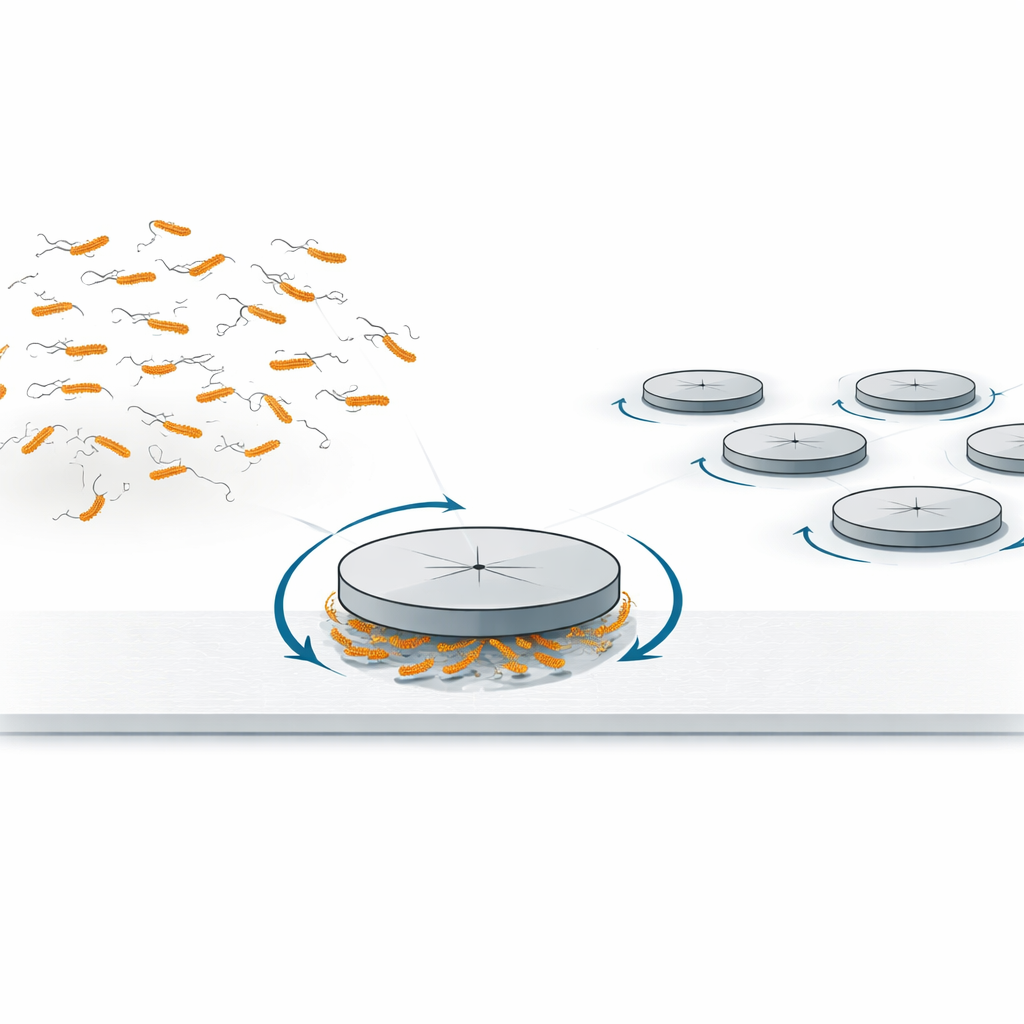
从随机碰撞到可控旋转
作者首先重新考察了一种更熟悉的效应:当光滑的圆盘(或“冰球”)放在充满浓密游动细菌的狭窄玻璃腔室底壁上时,细菌与圆盘边缘的碰撞使圆盘缓慢顺时针旋转。这一行为之前也在形状不规则的聚集体中观察到,其原因可归结为在固体表面附近,E. coli自然沿弯曲的顺时针路径游动。这些弯曲轨迹导致朝一个旋转方向的碰撞略多于另一方向,从而在圆盘周边产生微弱的净力矩。团队测量了旋转速率与圆盘尺寸的依赖关系,显示其标度符合这种边缘碰撞机制的预期,确认了与游动细菌的简单冲击可以使对称物体开始旋转。
在圆盘下方将单个细菌限制在通道内
为探测更微妙的运动来源,研究者使用高分辨率三维打印雕刻出在圆盘下方只有几微米高和宽的窄地下通道。在一种设计中,四个短径向腔室延伸至接近圆盘中心;在另一种设计中,一条直通道横贯圆盘,两端均开放。这些结构朝下开口,因此在底表面游动的细菌偶尔会进入并被紧密限制在冰球下方的通道中。由于通道非常狭窄,细菌不能轻易转身或沿墙滑过以简单地顶在死端上发力。然而,一旦单个细胞进入径向腔室,圆盘的旋转速率就跃升了一个数量级,始终以相同的(顺时针)方向旋转,且随着更多腔室被填满而进一步增加。即使当通道两端开放——因此没有墙可供顶靠——单个细菌穿过圆盘时也会在圆盘角度上产生特征性的“先下后上”变化:细胞通过时圆盘先朝一个方向转动,随后在细胞离开时向相反方向转动。关键是,这一模式与细菌是从左到右还是从右到左游动无关,从而排除了简单推进作为原因。
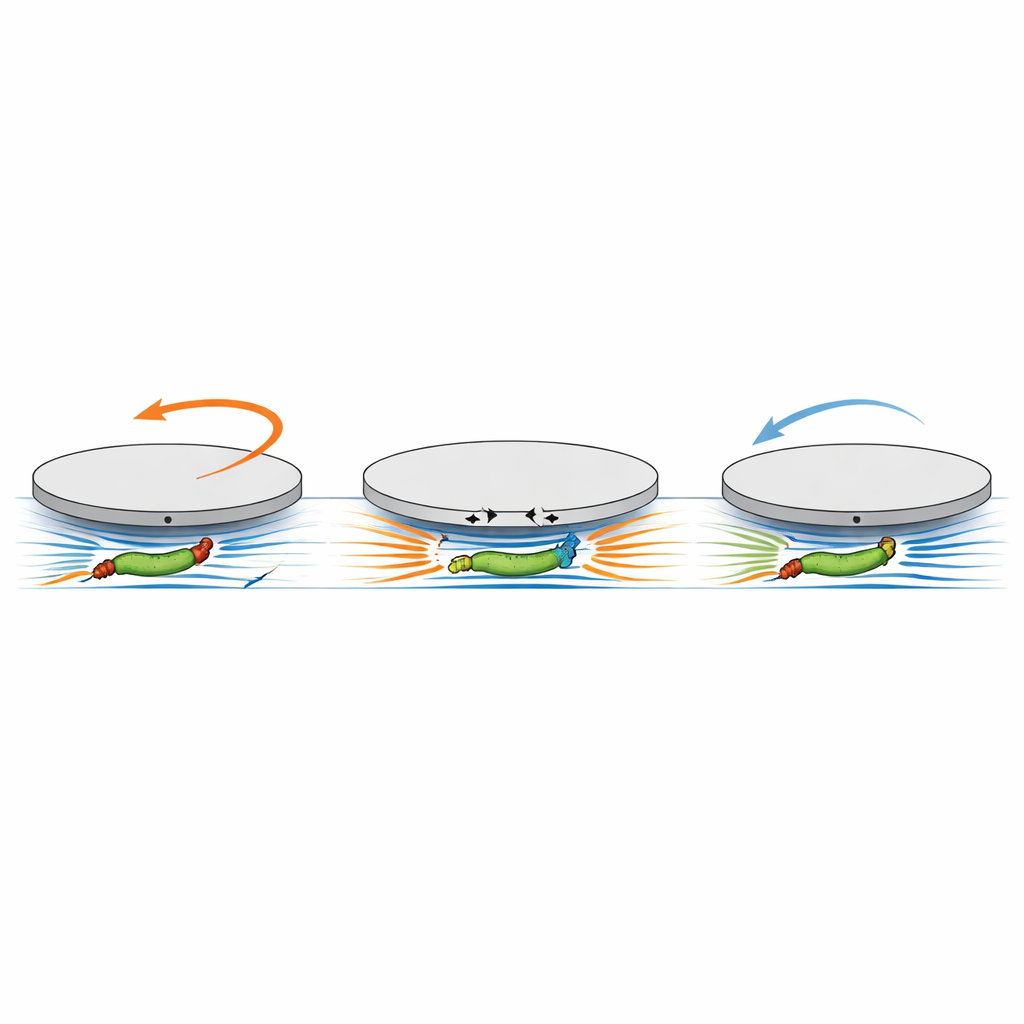
抓住圆盘的隐含力矩
为了解释这些令人费解的观测,团队建立了一个流体力学模型,关注的是细菌马达的扭转动作,而非直线推动。在模型中,旋转的细胞体和反向旋转的鞭毛束被视为流体中两个微小的旋转源,它们之间的间隔与细胞长度相当。当这对旋转源位于紧贴圆盘下方的窄通道内时,它们产生的旋转流在通道顶壁——圆盘的底面——的不同位置以相反方向施加牵引力。由于这两种牵引模式在通道方向上有所偏移,它们不会完全相互抵消。相反,它们结合起来产生一个倾向于使整个圆盘旋转的净力矩。计算显示,该力矩与细胞的游动方向无关,并且与体与鞭毛之间的有效间距成比例,而该间距又随细胞长度增长。模型既再现了当细胞体和鞭毛同时位于圆盘下时的初始顺时针转动,也再现了当细胞离开时仅剩一侧留在通道内导致的方向反转。
走向手性流体与微生物机器
通过将测量结果与模型比较,作者得出结论:E. coli的旋转马达充当了可以将扭转运动通过流体传递给附近物体的“力矩偶极”,而无需直接接触或形状不对称。限制性空间——在此为圆盘下的狭窄通道——将局部的扭转转化为持久的定向旋转。当许多这样的圆盘放置在细菌浴中时,它们可以形成一组全部朝同一方向旋转的相同转子,这是迈向“手性”流体的一步,此类流体的整体行为取决于总体的扭转方向。除了为由活细胞驱动的微观机器设计提供新途径外,该机制在细菌带有旋转鞭毛并穿过拥挤或多孔环境(如土壤、生物膜或工程过滤器)时也可能发挥作用,细微地将它们自身的导航与周围物体的运动耦合起来。
引用: Grober, D., Dhar, T., Saintillan, D. et al. The hydrodynamic torque dipole from rotary bacterial flagella powers symmetric discs. Nat. Phys. 22, 620–627 (2026). https://doi.org/10.1038/s41567-026-03189-4
关键词: 细菌运动性, 微流体学, 活性物质, 微观游动者, 微型机器人