Clear Sky Science · ru
Переосмысление филогении терапсид с помощью байесовских и кладистических подходов
Скрытая глава в нашей родословной
Млекопитающие, включая людей, ведут свое происхождение от древней группы «псевдомлекопитающих» — терапсидов, которые бродили по Земле задолго до динозавров. Однако ключевой отрезок их ранней истории отсутствует в палеонтологическом слое, что порождает большие вопросы о том, когда и как эти животные впервые диверсифицировались. В этом исследовании авторы возвращаются к этой загадке, применяя современные статистические инструменты, и показывают, что терапсиды, вероятно, возникли раньше, чем свидетельствуют их окаменелости, и быстро разветвились на главные линии, которые в конечном счете привели к млекопитающим.
Древние родственники в фрагментарном ископаемом ряду
Терапсиды были доминирующими наземными позвоночными в пермский период, примерно 300–250 миллионов лет назад. Они включают несколько крупных групп, некоторые из которых в конечном счете дали начало истинным млекопитающим. Но в отложениях, которые должны фиксировать их раннюю эволюцию, есть загадочный интервал с очень небольшим числом ископаемых, известный как пробел Олсона. До этого пробела мир заполнен более примитивными родственниками; после него внезапно появляются уже сформированные терапсидные группы во многих регионах мира. Такой резкий скачок затрудняет установление родственных связей между группами и оценку скорости их эволюции.
Применение новой математики к старым костям
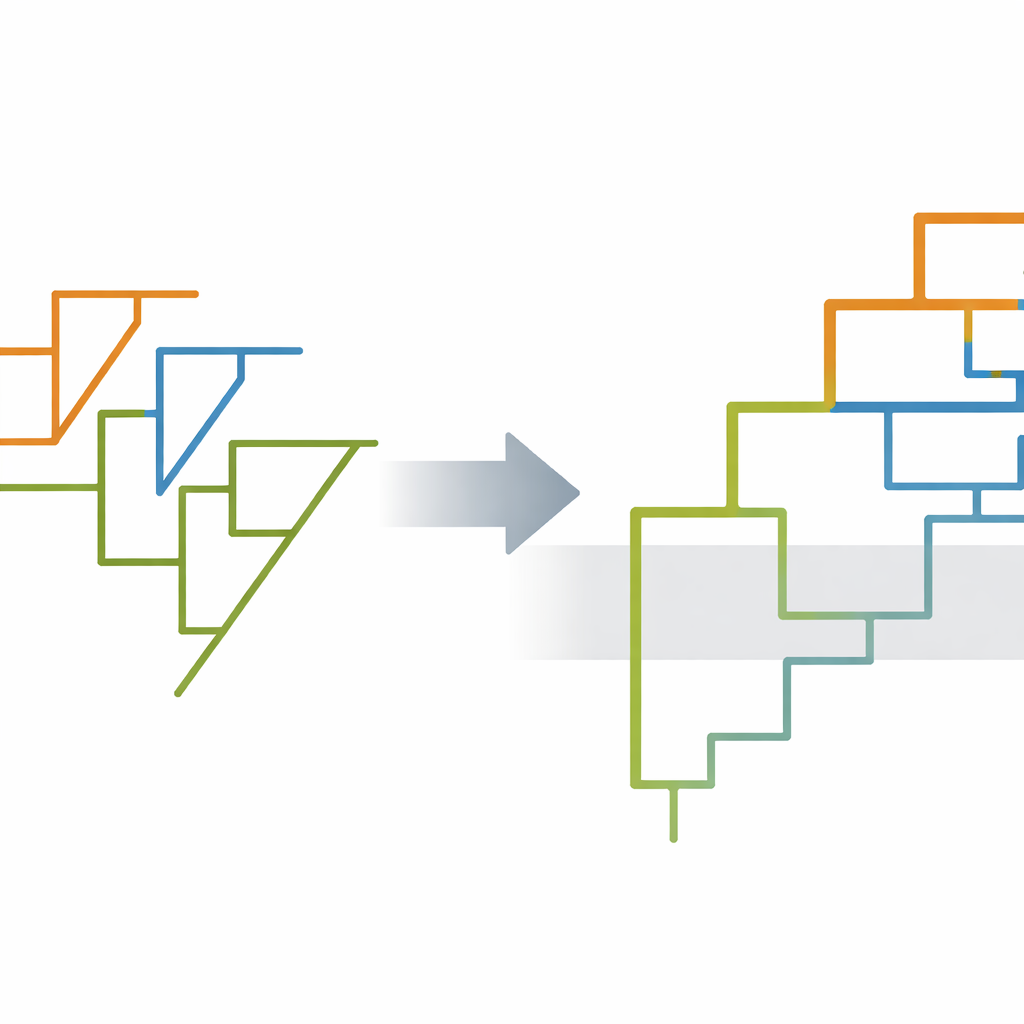
Большинство предыдущих исследований пытались восстановить отношения терапсидов, используя традиционный метод парсимонии, который ищет максимально простое эволюционное дерево. Авторы этой работы вместо этого собрали новый набор данных, сосредоточенный на черепных признаках 42 ранних видов терапсидов и их ближайших нетерапсидных родственников, закодировав 99 анатомических признаков. Затем они применили байесовские подходы — вероятностные методы, которые могут явно моделировать неопределенность и оценивать время расхождений — реализованные в двух программных пакетах, MrBayes и RevBayes. Анализы использовали модели, комбинирующие возрасты окаменелостей с процессами «рождение‑смерть», описывающими разветвление (видообразование), вымирание и окаменевание во времени.
Перестановка ветвей терапсидного древа
Байесовские деревья более детально разрешены по сравнению с результатами парсимонии и опровергают некоторые давние представления. Они поддерживают крупную подгруппу, названную Неотерапсиды (Neotherapsida), в которой растительноядные аномодонты выступают ближайшими родственниками преимущественно хищных териодонтов. Внутри териодонтов саблезубые горгонопсии выступают сестринской группой к кладе, включающей тероцефалий и цинодонтов, последние из которых в конечном итоге привели к млекопитающим. Исследование также обнаруживает свидетельства того, что две крупные группы, биармозухии (Biarmosuchia) и диноцефалии (Dinocephalia), образуют единый более широкий ответвление — связь, намечавшуюся в старых описательных работах, но не получавшую сильной поддержки в ранних компьютерных анализах. Несколько известных китайских видов из фауны Дашанкоу — Raranimus, Biseridens и Sinophoneus — смещаются на новые позиции у основания древа, ослабляя прежние утверждения о том, что некоторые из них принадлежат хорошо очерченным поздним линиям.
Заполнение временного пробела
Комбинируя анатомию с возрастами окаменелостей и моделью фоссилизованного процесса «рождение‑смерть», авторы оценили, когда основные терапсидные группы разошлись друг от друга. Их результаты указывают на происхождение линии терапсидов в раннем пермском периоде, до начала пробела Олсона, несмотря на отсутствие четких ископаемых этого времени. Большинство ныне известных крупных групп — такие как Biarmosuchia, Dinocephalia, Anomodontia, Gorgonopsia и Eutheriodontia —, по-видимому, быстро диверсифицировались между примерно 281 и 272 миллионами лет назад, перекрывая сам пробел. Дополнительное моделирование показывает, что имели место всплески повышенного вымирания, включая один во время пробела Олсона, тогда как скорости окаменевания оставались относительно стабильными. Эта картина согласуется со сценарием, в котором более раннее вымирание подрезало старые линии, одновременно открыв экологические ниши для диверсификации терапсидов.
Что это значит для восхождения млекопитающих
Для неспециалиста главный вывод в том, что наши предки‑млекопитающие не появились внезапно полностью сформированными и не ползли вперед в полной неизвестности. Вместо этого работа поддерживает двухэтапную историю: слабо задокументированная ранняя фаза, в ходе которой терапсиды постепенно отделялись от более рептильных предков, а затем быстрый взрыв диверсификации примерно в период пробела Олсона. Новые байесовские анализы реорганизуют ветви терапсидного древа и предлагают, что некоторые группы, считавшиеся отдельными, на самом деле могут образовывать более широкие линии, а цинодонты — прямые предшественники млекопитающих — вероятно, произошли из животных териоцефального уровня. По мере обнаружения новых окаменелостей, особенно из отложений, охватывающих пробел Олсона, эта уточненная схема поможет палеонтологам точнее их разместить и дальше прояснить, как и когда наши отдаленные предки сделали первые шаги к становлению млекопитающими.
Цитирование: Duhamel, A., Wynd, B., Wright, A.M. et al. Rethinking therapsid phylogeny through Bayesian and cladistic approaches. Sci Rep 16, 13171 (2026). https://doi.org/10.1038/s41598-026-38195-2
Ключевые слова: терапсиды, пермские ископаемые, филогения, байесовская эволюция, пробел Олсона