Clear Sky Science · ru
Энторинальная кора кодирует удалённые релевантные для задачи локации независимо от CA1
Как мозг связывает места во время пауз
Представьте, что вы стоите у входной двери и представляете кухню, где оставили кофе. Наш мозг постоянно перескакивает между тем, где мы находимся, и тем, куда нам нужно попасть. В этом исследовании поставили вопрос, как ключевая область навигации у мыши — медиальная энторинальная кора — обрабатывает такие мысленные «перепрыгивания» между локациями, когда животное неподвижно, и что это может значить для того, как мы усваиваем маршруты и цели.
Лабиринт, проверяющий память о месте
Чтобы изучить это, исследователи обучали мышей в лабиринте в форме буквы X, где в каждом испытании были связаны две точки с вознаграждением. Сначала мышь посещала «образцовую» ветвь, чтобы получить небольшое лакомство, затем выбирала между двумя «выборными» ветвями, чтобы заработать большее вознаграждение по правилу: перейти в ветвь на той же стороне лабиринта, а после смены правила — в ветвь на противоположной стороне. В течение многих дней мыши усваивали эти меняющиеся правила, выполняя около сотни испытаний за сессию. Пока они выполняли задачу, ультратонкие зонды Neuropixels записывали активность сотен отдельных нейронов в медиальной энторинальной коре и в соседней области гиппокампа CA1, что позволило декодировать, какие места лабиринта каждая популяция клеток представляла в каждый момент времени.
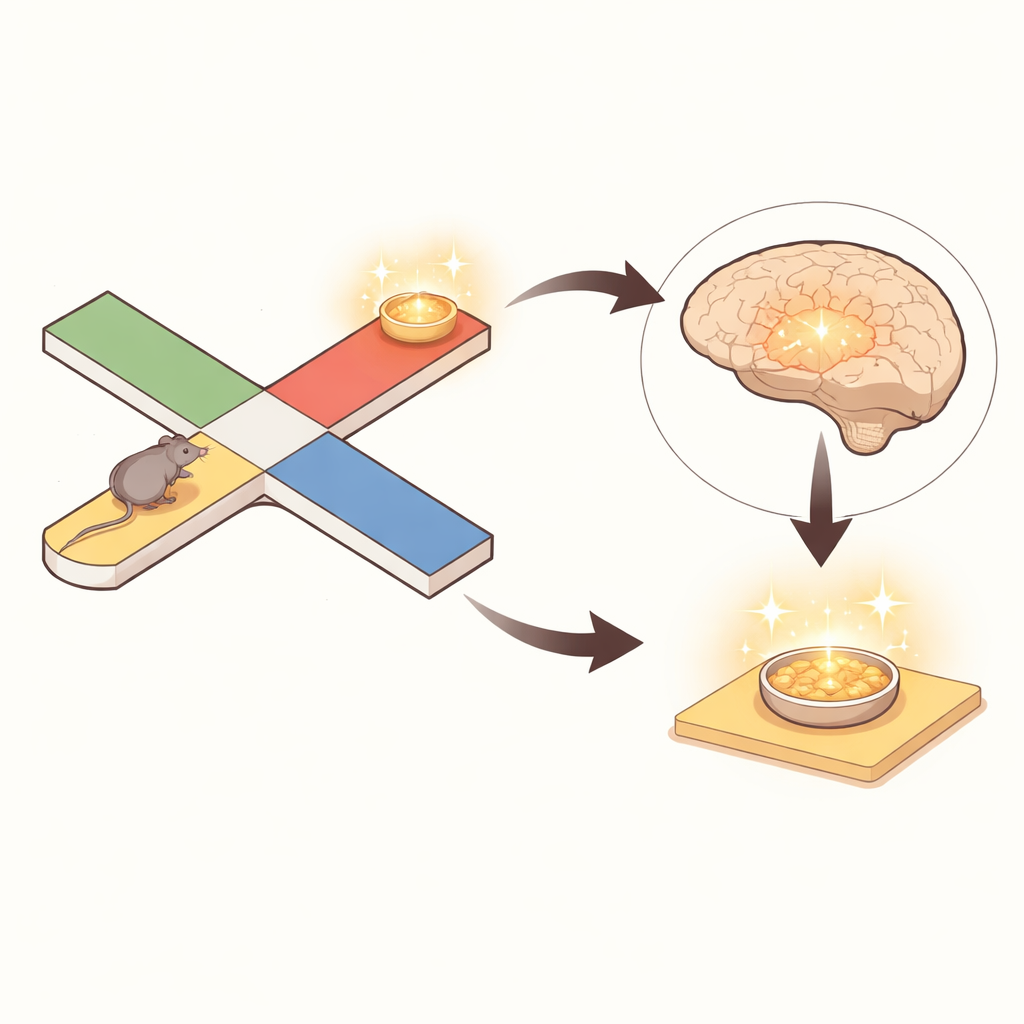
Карта мозга «перепрыгивает» в моменты неподвижности
Во время движения паттерны активности в энторинальной коре тесно соответствовали реальному положению мыши в лабиринте, подобно движущейся точке на GPS‑экране. Но когда мышь делала паузу, происходило нечто поразительное: декодированная позиция по активности энторинальной коры часто «перепрыгивала» к локациям, удалённым от тела животного, нередко — на противоположную сторону лабиринта. Авторы определяли такие перепрыжки как «нелокальное» кодирование, если представляемая точка находилась как минимум в 20 сантиметрах. Почти в половине всех пауз встречался такой нелокальный контент, а примерно в четверти всех временных отрезков покоя отражались удалённые позиции. Важно, что это не была ошибка декодирования. Клетки, настроенные на текущую физическую позицию, продолжали фиpировать, но клетки, предпочитающие удалённые места в лабиринте, усиливали активность в эти эпизоды и сдвигали чтение в сторону тех далеких позиций.
Независимые «снимки», а не классическое воспроизведение
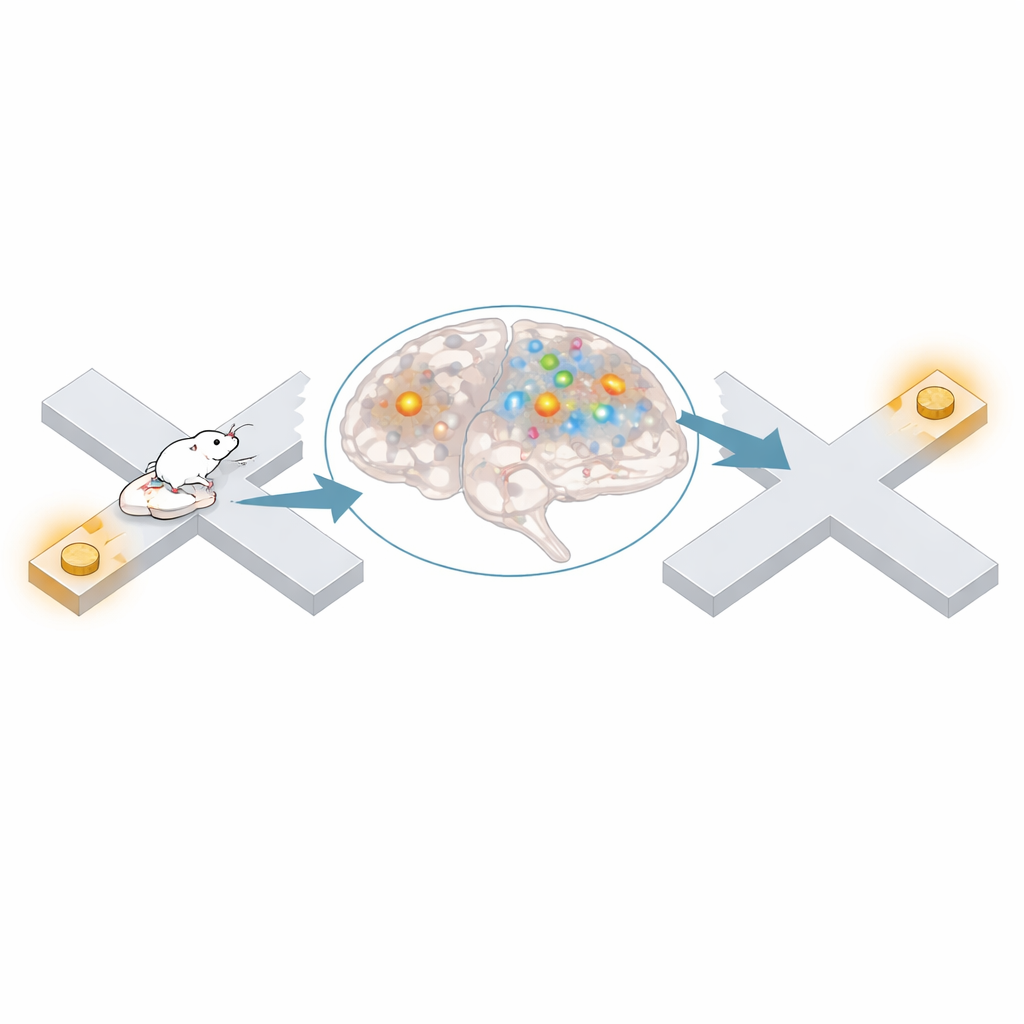
Ранее показывали скоординированное «воспроизведение» пройденных маршрутов во время кратких электрических событий в гиппокампе, называемых sharp‑wave ripples, которые, как полагают, поддерживают память. Здесь энторинальная кора иногда отображала дальние позиции во время этих рипплов, но большая часть нелокального кодирования происходила вне них. При сравнении активности энторинальной коры и CA1 выяснилось, что две области были менее синхронизованы в периоды нелокальности, чем в локальные периоды. CA1 реже представлял ту же локацию, что и энторинальная кора, пары клеток между областями реже фиpировали совместно, и быстрый ритм, который, как полагают, передаёт энторинальный вход в CA1, был слабее. Эти результаты указывают на то, что во многих таких мысленных перепрыжках энторинальная кора ведёт собственную внутреннюю карту с ограниченным влиянием на выход гиппокампа.
Думать о нужном месте в нужный момент
Содержание этих удалённых представлений было далеко не случайным. Мыши склонны дольше задерживаться у точек с вознаграждением, но нелокальное кодирование особенно часто возникало, когда они находились в других частях лабиринта, например в центральной ветви. Независимо от того, где физически находилось животное, удалённые представления предпочтительнее отражали места с вознаграждением гораздо чаще, чем случайно. Когда мышь задерживалась у образцовой награды, активность энторинальной коры чаще представляла ту выборную награду, которая в успешных испытаниях была правильно связана с этим образцом, а при ошибках сдвигалась к несвязанной награде. Аналогично, отдыхая у выборной награды после правильного решения, энторинальная карта чаще «перепрыгивала» назад к соответствующей образцовой награде. При смене правила задачи, когда «правильная» пара ветвей менялась, предпочтительные удалённые представления также менялись в соответствии с новым правилом.
Почему это важно для повседневной навигации
Эти результаты показывают, что медиальная энторинальная кора часто представляет релевантные для задачи, но физически удалённые места, когда животное неподвижно, и что эти мысленные перепрыжки настроены на правильные парные связи между локациями. Пока другая область мозга, CA1, в основном отключается в эти моменты, энторинальная кора может тихо усиливать связи между связанными точками или помогать животному вызвать полезную цель при решении, куда идти дальше. В повседневных терминах, когда вы делаете паузу у одного ориентира и вспоминаете другой, это исследование предлагает, что внутренняя карта вашего мозга может активно связывать эти места в фоновом режиме, даже когда вы не движетесь.
Цитирование: Aery Jones, E.A., Low, I.I.C., Cho, F.S. et al. Entorhinal cortex represents task-relevant remote locations independently of CA1. Nat Neurosci 29, 1181–1190 (2026). https://doi.org/10.1038/s41593-026-02232-0
Ключевые слова: пространственная навигация, энторинальная кора, воспроизведение памяти, лабиринт для мышей, гиппокамп