Clear Sky Science · pl
Kora śródwęchowa reprezentuje odległe, istotne dla zadania miejsca niezależnie od CA1
Jak mózg łączy miejsca, gdy się zatrzymujemy
Wyobraź sobie, że stoisz w drzwiach i wyobrażasz sobie kuchnię, w której zostawiłeś kawę. Nasze mózgi ciągle przeskakują między tym, gdzie jesteśmy, a gdzie musimy dotrzeć. W tym badaniu pytano, jak kluczowy obszar nawigacyjny w mózgu myszy — przyśrodkowa kora śródwęchowa — radzi sobie z takimi mentalnymi przeskokami między miejscami, gdy zwierzę stoi w miejscu, i co to może znaczyć dla uczenia się tras i celów.
Labirynt testujący pamięć miejsc
Aby to zbadać, badacze wyszkolili myszy w labiryncie w kształcie X, gdzie każdy trial łączył dwa nagradzane miejsca. Mysz najpierw odwiedzała ramię „próbkowe”, by otrzymać małą nagrodę, a następnie wybierała między dwoma ramionami „wyboru”, aby zdobyć większą nagrodę według reguły: najpierw iść do ramienia po tej samej stronie labiryntu, a po odwróceniu reguły — do ramienia po przeciwnej stronie. Przez wiele dni myszy uczyły się tych zmieniających się zasad, wykonując około stu prób w sesji. W trakcie zadania ultracienkie sondy Neuropixels rejestrowały aktywność setek pojedynczych komórek nerwowych w przyśrodkowej korze śródwęchowej i w sąsiednim regionie, hipokampie CA1, co pozwoliło zespołowi odkodować, które miejsca w labiryncie reprezentowała każda z populacji komórek w danym momencie.
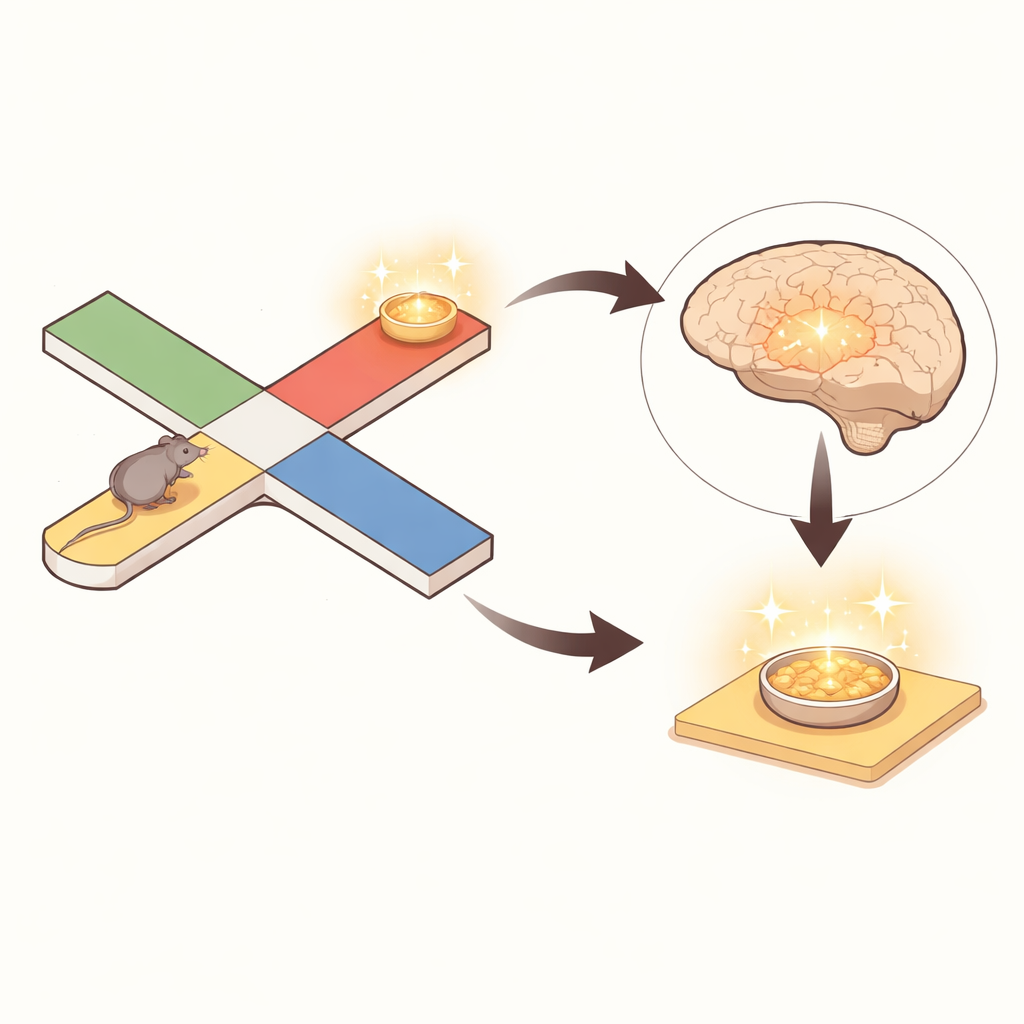
Mapa mózgu przeskakuje podczas chwilowego bezruchu
Podczas ruchu wzorce aktywności w korze śródwęchowej ściśle śledziły rzeczywistą pozycję myszy w labiryncie, niczym poruszający się punkt na wyświetlaczu GPS. Gdy jednak mysz się zatrzymywała, działo się coś uderzającego: odkodowana pozycja z aktywności śródwęchowej często „przeskakiwała” do miejsc odległych od ciała zwierzęcia, często na przeciwległą stronę labiryntu. Autorzy zdefiniowali te przeskoki jako kodowanie „nielokalne”, gdy reprezentowane miejsce znajdowało się co najmniej 20 centymetrów dalej. Prawie połowa wszystkich zatrzymań zawierała takie nielokalne treści, a około jedna czwarta wszystkich przedziałów czasu spędzonych w bezruchu odzwierciedlała odległe miejsca. Co ważne, nie był to błąd dekodowania. Komórki dostrojone do bieżącej fizycznej lokalizacji nadal aktywowały się, ale komórki, których preferowane miejsca leżały w odległych częściach labiryntu, zwiększały swoją aktywność podczas tych epizodów i przesuwały odczyt w kierunku tych dalekich pozycji.
Niezależne migawki, a nie klasyczny replay
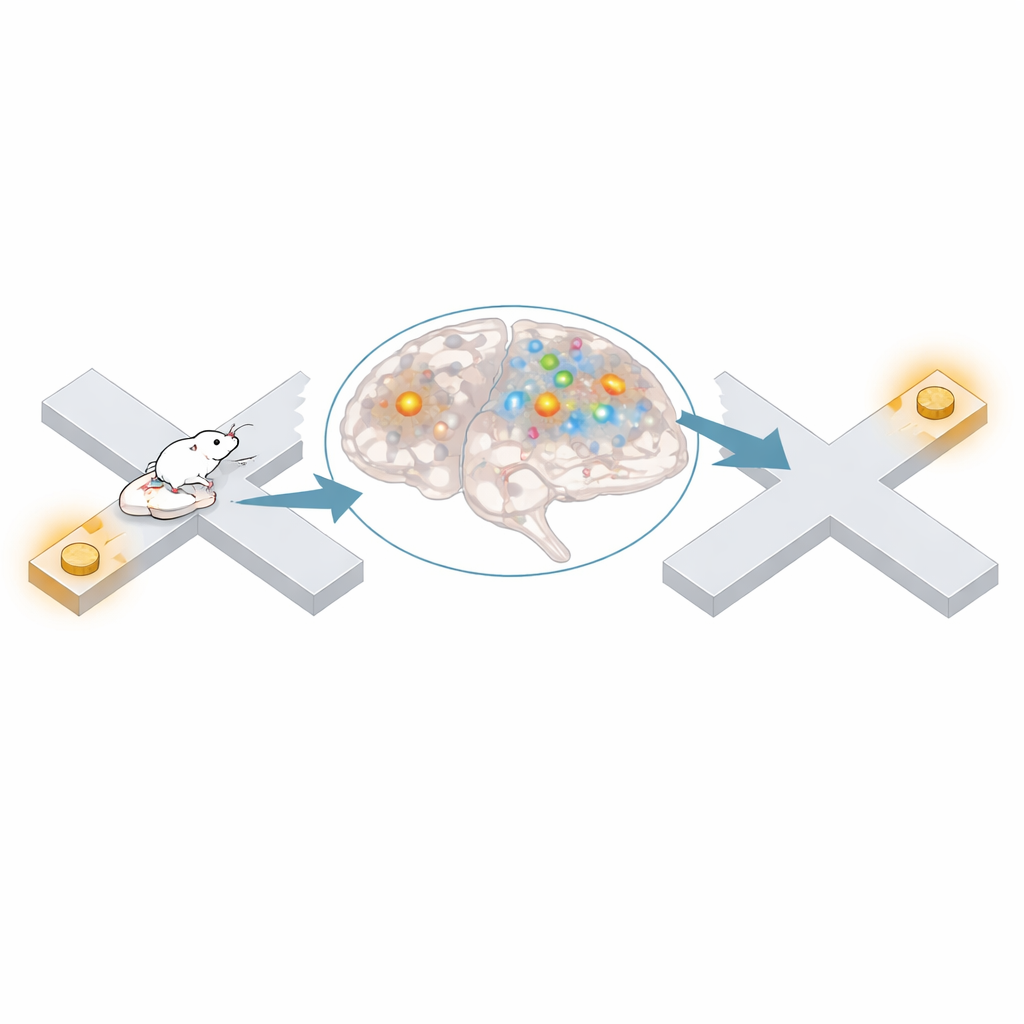
Poprzednie prace wykazały skoordynowany „replay” przebytych ścieżek podczas krótkich zjawisk elektrycznych w hipokampie zwanych sharp-wave ripple, które uważa się za wspierające pamięć. Tutaj kora śródwęchowa czasami reprezentowała odległe pozycje podczas tych ripple, ale większość nielokalnego kodowania występowała poza nimi. Porównując aktywność śródwęchową i CA1, zespół stwierdził, że oba regiony były mniej zsynchronizowane podczas okresów nielokalnych niż podczas lokalnych. CA1 rzadziej reprezentował tę samą lokalizację co kora śródwęchowa, pary komórek między regionami rzadziej wyładowywały się razem, a szybkie rytmy uznawane za przenoszące wejście śródwęchowe do CA1 były słabsze. Wyniki te sugerują, że podczas wielu z tych mentalnych przeskoków kora śródwęchowa prowadzi własną wewnętrzną mapę z ograniczonym wpływem na wyjście hipokampa.
Myślenie o właściwym miejscu we właściwym czasie
Treść tych odległych reprezentacji była daleka od losowej. Myszy miały tendencję do dłuższego zatrzymywania się przy miejscach z nagrodą, ale nielokalne kodowanie było szczególnie prawdopodobne, gdy znajdowały się w innych częściach labiryntu, na przykład w ramieniu centralnym. Niezależnie od tego, gdzie zwierzę rzeczywiście siedziało, odległe reprezentacje faworyzowały miejsca nagradzane znacznie częściej niż przypadkowo. Gdy mysz zatrzymywała się przy nagrodzie próbkowej, aktywność śródwęchowa częściej reprezentowała nagrodę wyboru powiązaną z tą próbką w poprawnych próbach, a w błędach przesuwała się w stronę niepowiązanej nagrody. Podobnie, odpoczywając przy nagrodzie wyboru po poprawnej decyzji, mapa śródwęchowa częściej przeskakiwała z powrotem do odpowiadającej nagrody próbki. Gdy reguła zadania została odwrócona tak, że „poprawne” powiązanie między ramionami się zmieniło, preferowane odległe reprezentacje zmieniały się równolegle do nowej reguły.
Dlaczego to ma znaczenie dla codziennej nawigacji
Wyniki te wskazują, że przyśrodkowa kora śródwęchowa często reprezentuje istotne dla zadania, lecz fizycznie odległe miejsca, gdy zwierzę stoi w miejscu, i że te mentalne przeskoki są dostrojone do poprawnych powiązań między lokalizacjami. Podczas gdy inny obszar mózgu, CA1, w dużej mierze się wyłącza w tych epizodach, kora śródwęchowa może cicho wzmacniać powiązania między sparowanymi punktami lub pomagać zwierzęciu przywołać użyteczny cel, gdy decyduje, dokąd pójść dalej. W codziennych kategoriach, gdy zatrzymujesz się przy jednym punkcie orientacyjnym i przypominasz sobie inny, to badanie sugeruje, że wewnętrzna mapa twojego mózgu może aktywnie łączyć te miejsca w tle, nawet gdy się nie poruszasz.
Cytowanie: Aery Jones, E.A., Low, I.I.C., Cho, F.S. et al. Entorhinal cortex represents task-relevant remote locations independently of CA1. Nat Neurosci 29, 1181–1190 (2026). https://doi.org/10.1038/s41593-026-02232-0
Słowa kluczowe: nawigacja przestrzenna, kora śródwęchowa, replay pamięci, labirynt myszy, hipokamp