Clear Sky Science · ru
Ось PI(3,5)P2/CHMP4B на лизосомах необходима для микроаутофагического разрушения STING
Как клетки успокаивают внутреннюю тревогу
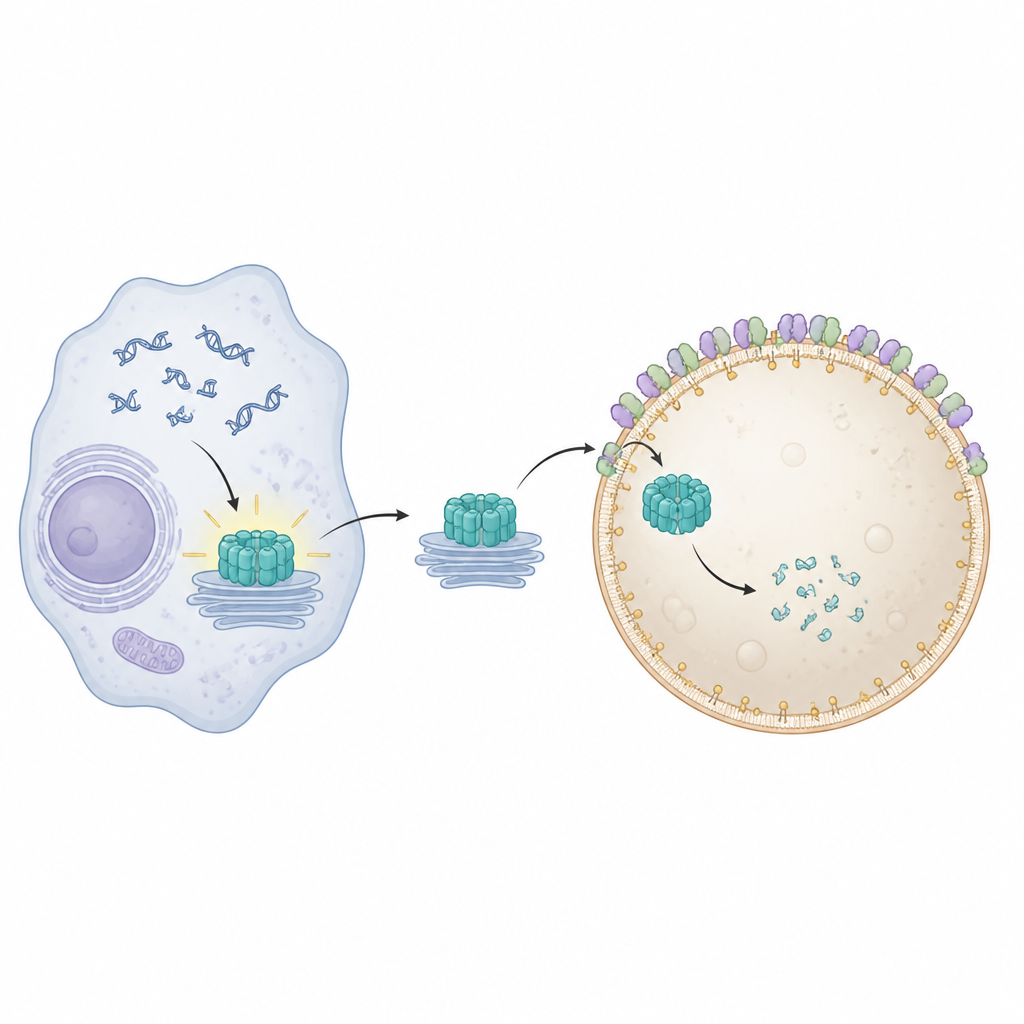
В наших клетках есть внутренняя система тревоги, которая замечает попавшую в неположенное место ДНК — признак вирусной инфекции или повреждения клетки. Ключевой переключатель этой тревоги, называемый STING, помогает мобилизовать противовирусные и провоспалительные ответы. Но, как и любую сигнализацию, STING нужно выключать после устранения угрозы, иначе это может привести к хроническому воспалению и болезни. В этом исследовании показано, как крошечные компоненты мембраны на центрах утилизации клетки — лизосомах — работают совместно, чтобы «проглотить» и разрушить STING, возвращая клетке спокойствие.
ДНК‑тревога клетки и связанные с ней риски
STING располагается на внутренних мембранах и активируется при обнаружении ДНК в неправильном месте внутри клетки. После активации он перемещается со своего обычного участка у ядра через несколько мембранных станций и запускает выработку противовирусных и провоспалительных молекул. Чтобы избежать длительного повреждения, клетки должны быстро убирать отслуживший STING. Предыдущие работы показали, что специальные белки, перестраивающие мембраны и объединённые под названием ESCRT, помогают лизосомам отсекать и поглощать груз прямо с их поверхности в процессе, называемом микроаутофагией. Однако оставалось неясным, как именно STING физически оборачивается мембраной и попадает внутрь лизосом.
Поиск молекулярного выключателя
Исследователи сначала разработали чувствительный тест, отслеживающий скорость разрушения STING в клетках, прикрепив к нему светопродуцирующие ферменты. Затем они применили панель ингибиторов киназ — препаратов, блокирующих различные сигнальные ферменты — чтобы выявить те, которые замедляют деградацию STING. Несколько соединений выделялись, особенно те, что блокировали фермент Pikfyve. Pikfyve участвует в создании редкого сигнального липида PI(3,5)P2 на поздних компартментах клетки, включая лизосомы. Ингибирование Pikfyve снижало уровень PI(3,5)P2, мешало эффективной деградации STING и приводило к стойкой активации сигналов STING, что удлиняло экспрессию провоспалительных генов в иммунных клетках мыши и человека.
Наблюдение за тем, как STING застревает у внешней стороны утилизационного центра клетки
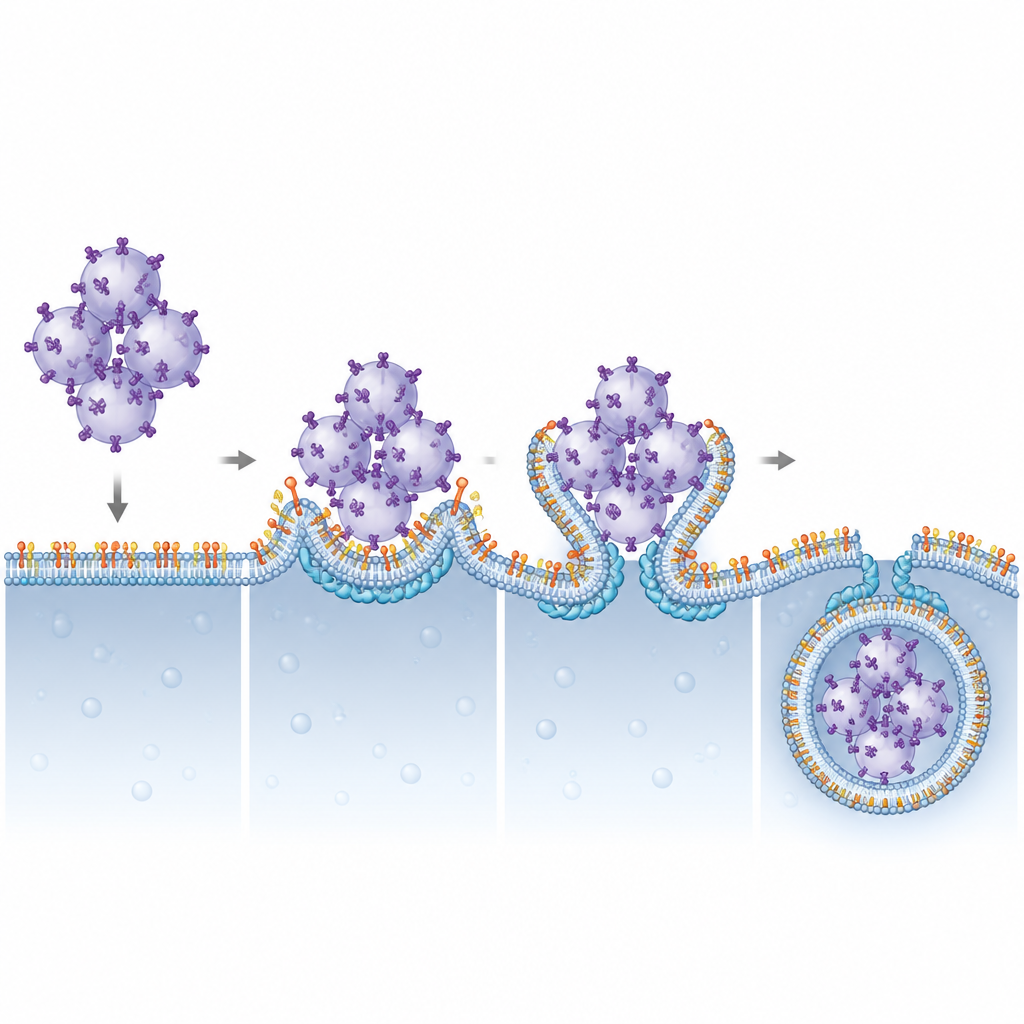
Чтобы визуализировать, что именно шло не так при блокаде Pikfyve, команда использовала продвинутую флуоресцентную и электронную микроскопию. В норме активированный STING перемещается к эндосомам‑переработчикам, упаковывается в кластеры маленьких везикул, и эти кластеры затем проглатываются лизосомами. При ингибировании Pikfyve лизосомы увеличивались в размере, но не могли поглотить кластеры везикул, богатых STING. Вместо этого сотни мелких везикул со STING накапливались прямо снаружи лизосом, хотя STING всё ещё нес на себе молекулярные метки, которые обычно помечают его для утилизации. Это показало, что проблема заключалась не в распознавании STING как «мусора», а на финальном этапе, где лизосомы огибают и внутренне захватывают груз.
Пара липида и белка, которая обеспечивает «проглатывание» мембраны
Далее авторы спросили, какие компоненты ESCRT зависят от PI(3,5)P2 для работы на лизосомах. Систематически уменьшая экспрессию различных субъединиц ESCRT, они выделили CHMP4B, часть группы ESCRT‑III, как критически важный для разрушения STING. CHMP4B обычно образует динамичные филаменты, способные стягивать и перерезать мембраны. Визуализация показала, что CHMP4B локализуется на мембранах лизосом, и эта локализация исчезает при блокаде Pikfyve или синтеза PI(3,5)P2. Компьютерные моделирования и биохимические тесты выявили, что небольшой кластер положительно заряженных аминокислот на CHMP4B специфически распознаёт и связывает PI(3,5)P2. Мутации в этом кластере не позволяли CHMP4B прикрепляться к лизосомам, нарушали его связывание с PI(3,5)P2 и не восстанавливали деградацию STING или отключение сигнализации в клетках без нормального CHMP4B.
Почему это важно для иммунитета и болезней
Эта работа очерчивает ясное структурное и функциональное партнёрство между редким липидом мембраны PI(3,5)P2 и белком ESCRT‑III CHMP4B в выключении тревоги STING. Закрепляя CHMP4B на лизосомах, PI(3,5)P2 позволяет этим органеллам изгибать свои мембраны вокруг кластеров везикул, загруженных STING, и втягивать их внутрь для разрушения. Когда эта система нарушена, сигнализация STING сохраняется, что может объяснять воспалительные черты при расстройствах, связанных с функцией Pikfyve, и указывает на то, что точная настройка этой липид‑белковой оси может в будущем помочь усилить противоопухолевый иммунитет или сдержать вредное воспаление.
Цитирование: Shoji, T., Shinojima, A., Kishimoto, T. et al. A PI(3,5)P2/CHMP4B axis on lysosomes is essential for microautophagic degradation of STING. Nat Commun 17, 4602 (2026). https://doi.org/10.1038/s41467-026-72828-4
Ключевые слова: STING, лизосомальная микроаутофагия, PI(3,5)P2, ESCRT-III, врожденный иммунитет