Clear Sky Science · pl
Oś PI(3,5)P2/CHMP4B na lizosomach jest niezbędna do mikrofagicznego degradowania STING
Jak komórki uciszają wewnętrzny alarm
Nasze komórki mają wewnętrzny system alarmowy wykrywający obce DNA — sygnał zagrożenia wskazujący na zakażenie wirusowe lub uszkodzenie komórki. Kluczowy przełącznik alarmowy zwany STING pomaga uruchomić obronę przeciwwirusową i zapalną. Jednak jak każdy alarm, STING musi zostać wyłączony po opanowaniu zagrożenia, inaczej może prowokować przewlekłe zapalenie i choroby. Badanie to ujawnia, jak drobne składniki błonowe na komórkowych centrach recyklingu zwanych lizosomami współdziałają, by pochłonąć i rozłożyć STING, przywracając spokój w komórce.
Alarm DNA w komórce i jego zagrożenia
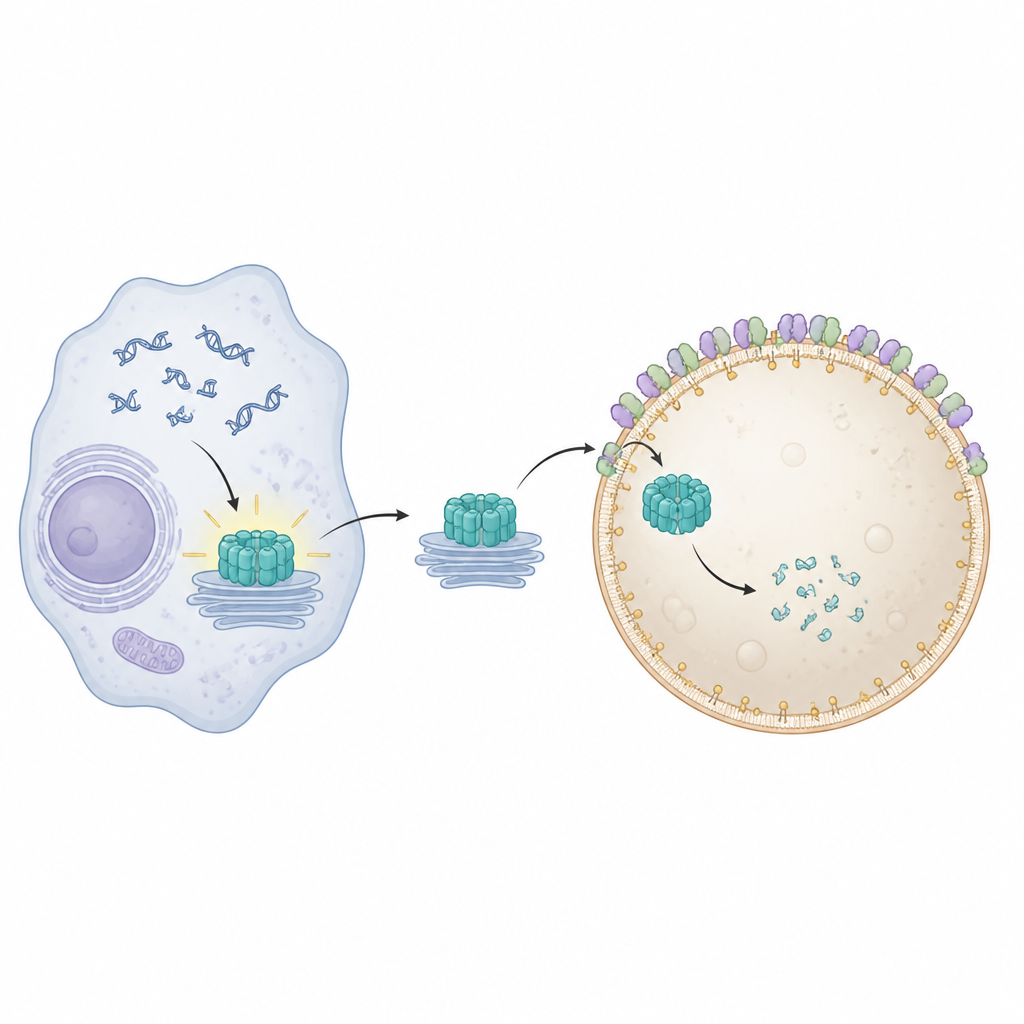
STING znajduje się na błonach wewnętrznych i aktywuje się, gdy wyczuje DNA w niewłaściwym miejscu w komórce. Po aktywacji przemieszcza się ze swojego zwykłego miejsca w pobliżu jądra przez kilka stacji błonowych i uruchamia produkcję cząsteczek przeciwwirusowych i zapalnych. Aby uniknąć trwałych szkód, komórki muszą szybko usuwać zużyty STING. Wcześniejsze prace pokazały, że specjalne białka remodelujące błony, znane łącznie jako ESCRT, pomagają lizosomom odcinać i pochłaniać ładunki bezpośrednio z ich powierzchni w procesie zwanym mikrofagią. Dokładny sposób, w jaki STING jest fizycznie owinięty i wchłaniany przez lizosomy, pozostawał jednak niejasny.
Odnalezienie molekularnego wyłącznika
Naukowcy najpierw opracowali czuły test śledzący, jak szybko komórki rozkładają STING, łącząc go z enzymami wytwarzającymi światło. Potem zastosowali panel inhibitorów kinaz, leków blokujących różne enzymy sygnalizacyjne, aby sprawdzić, które spowalniają rozkład STING. Kilka związków wyróżniało się, zwłaszcza te blokujące enzym o nazwie Pikfyve. Pikfyve uczestniczy w tworzeniu rzadkiego lipidu sygnałowego PI(3,5)P2 na późnych przegrodach komórkowych, w tym na lizosomach. Hamowanie Pikfyve zmniejszało poziom PI(3,5)P2, uniemożliwiało efektywną degradację STING i powodowało utrzymywanie się aktywowanych sygnałów STING, co prowadziło do przedłużonej ekspresji genów zapalnych zarówno w komórkach odpornościowych myszy, jak i ludzi.
Obserwacja, jak STING utknął poza komórkowym recyklerem
Aby zobrazować, co poszło nie tak po zablokowaniu Pikfyve, zespół użył zaawansowanej mikroskopii fluorescencyjnej i elektronowej. W normalnych warunkach aktywowany STING trafia do endosomów recyklingowych, pakuje się w skupiska małych pęcherzyków, które następnie są połykane przez lizosomy. Gdy Pikfyve był zablokowany, lizosomy powiększały się, ale nie potrafiły pochłonąć tych pęcherzykowych skupisk bogatych w STING. Zamiast tego setki małych pęcherzyków zawierających STING nagromadziły się tuż poza lizosomami, chociaż STING wciąż nosił molekularne znaczniki zwykle kwalifikujące go do utylizacji. To pokazało, że problem nie polegał na rozpoznaniu STING jako odpadu, lecz na końcowym etapie, w którym lizosomy owijają i internalizują ładunek.
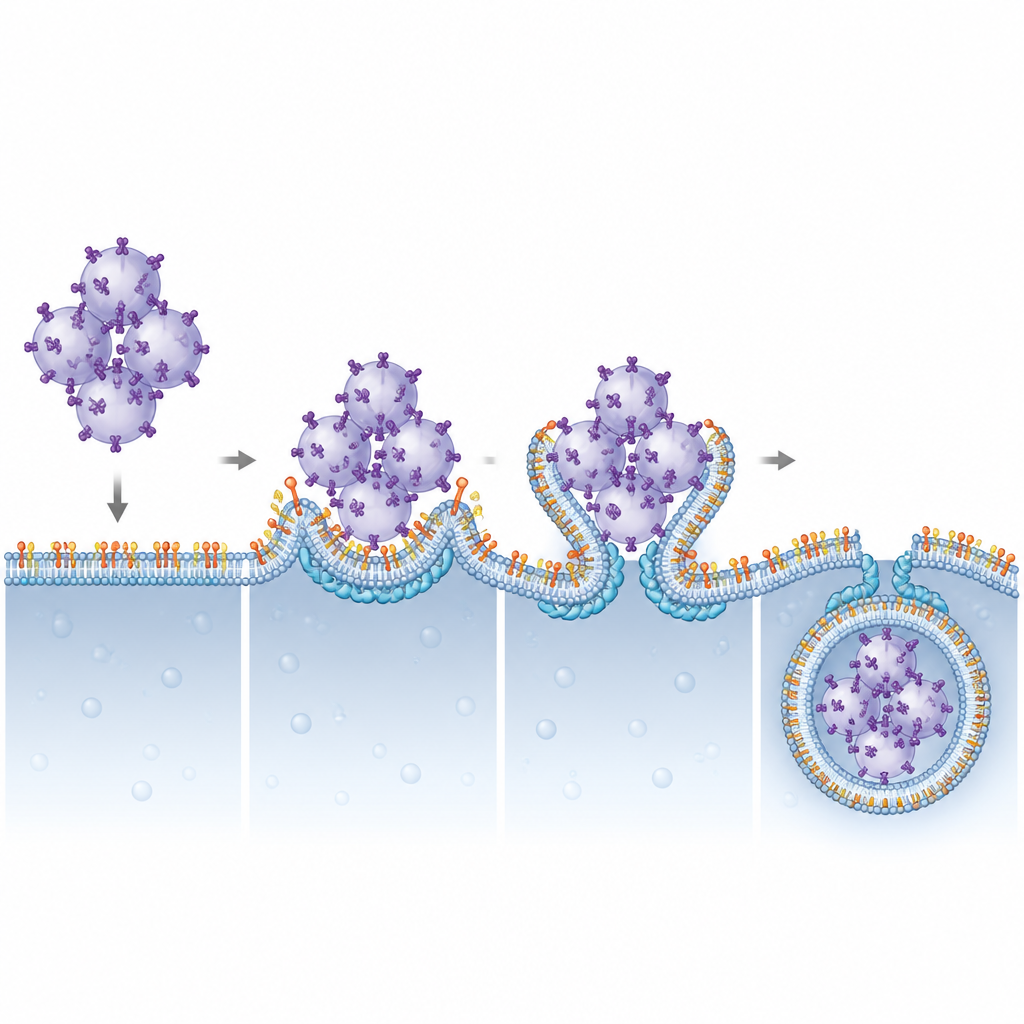
Para lipid–białko napędzająca połykanie błony
Następnie autorzy zapytali, które składniki ESCRT zależą od PI(3,5)P2, by działać na lizosomach. Poprzez systematyczne zmniejszanie różnych podjednostek ESCRT wskazali CHMP4B, należący do grupy ESCRT-III, jako kluczowy dla degradowania STING. CHMP4B zwykle tworzy dynamiczne filamenty, które potrafią obkurczać i przecinać błony. Obrazowanie wykazało, że CHMP4B lokalizuje się na błonach lizosomów, a ta lokalizacja znika, gdy blokowana jest produkcja Pikfyve lub PI(3,5)P2. Symulacje komputerowe i testy biochemiczne ujawniły, że mała grupa dodatnio naładowanych aminokwasów na CHMP4B specyficznie rozpoznaje i wiąże PI(3,5)P2. Mutacje w tej grupie uniemożliwiły CHMP4B przyłączanie się do lizosomów, zapobiegły wiązaniu PI(3,5)P2 i nie przywróciły degradacji STING ani wyciszenia sygnalizacji w komórkach pozbawionych normalnego CHMP4B.
Dlaczego to ma znaczenie dla odporności i chorób
Ta praca opisuje jasne strukturalne i funkcjonalne partnerstwo między rzadkim lipidm błonowym PI(3,5)P2 a białkiem ESCRT-III CHMP4B w wyłączaniu alarmu STING. Poprzez zakotwiczenie CHMP4B na lizosomach, PI(3,5)P2 pozwala tym organellom wyginać swoje błony wokół skupisk pęcherzyków z załadowanym STING i odcinać je do wnętrza w celu zniszczenia. Gdy ten system jest zaburzony, sygnalizacja STING utrzymuje się, co może tłumaczyć zapalne cechy w schorzeniach związanych z funkcją Pikfyve i sugeruje, że precyzyjne regulowanie tej osi lipid–białko może w przyszłości pomóc wzmocnić odpowiedź przeciwnowotworową lub ograniczyć szkodliwe zapalenie.
Cytowanie: Shoji, T., Shinojima, A., Kishimoto, T. et al. A PI(3,5)P2/CHMP4B axis on lysosomes is essential for microautophagic degradation of STING. Nat Commun 17, 4602 (2026). https://doi.org/10.1038/s41467-026-72828-4
Słowa kluczowe: STING, lizosomalna mikrofagia, PI(3,5)P2, ESCRT-III, wrodzona odporność