Clear Sky Science · en
A PI(3,5)P2/CHMP4B axis on lysosomes is essential for microautophagic degradation of STING
How Cells Calm an Internal Alarm
Our cells carry an internal alarm system that spots stray DNA, a danger sign of viral infection or cellular damage. A key alarm switch called STING helps summon antiviral and inflammatory defenses. But like any alarm, STING must be shut off once the threat is handled, or it can drive chronic inflammation and disease. This study reveals how tiny membrane components on cellular recycling centers called lysosomes work together to swallow and dismantle STING, restoring cellular calm.
The Cell’s DNA Alarm and Its Risks
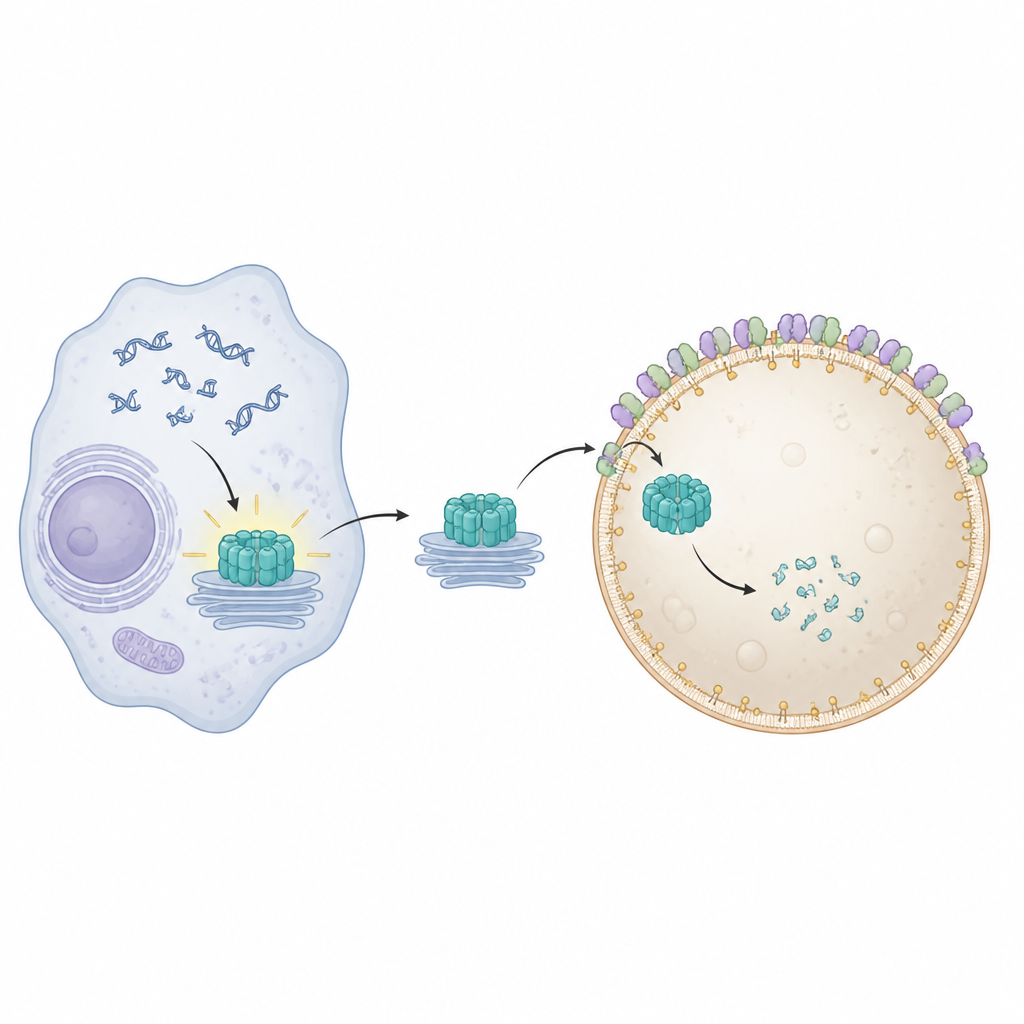
STING sits on internal membranes and turns on when it senses DNA in the wrong place inside a cell. Once activated, it moves from its usual home near the nucleus through several membrane stations and ignites production of antiviral and inflammatory molecules. To avoid lasting damage, cells must clear away used STING promptly. Earlier work showed that special membrane remodeling proteins, known collectively as ESCRT, help lysosomes pinch off and engulf cargo directly from their surface in a process called microautophagy. Exactly how STING becomes physically wrapped and taken in by lysosomes, however, has been unclear.
Finding a Molecular Off Switch
The researchers first built a sensitive test that tracks how quickly cells break down STING by attaching it to light-producing enzymes. They then applied a panel of kinase inhibitors, drugs that block many different signaling enzymes, to see which ones slowed STING breakdown. Several compounds stood out, particularly those that blocked an enzyme called Pikfyve. Pikfyve helps build a rare signaling fat called PI(3,5)P2 on late cellular compartments, including lysosomes. Inhibiting Pikfyve reduced PI(3,5)P2, prevented efficient STING degradation, and caused activated STING signals to linger, leading to prolonged expression of inflammatory genes in both mouse and human immune cells.
Watching STING Get Stuck Outside the Cell’s Recycler
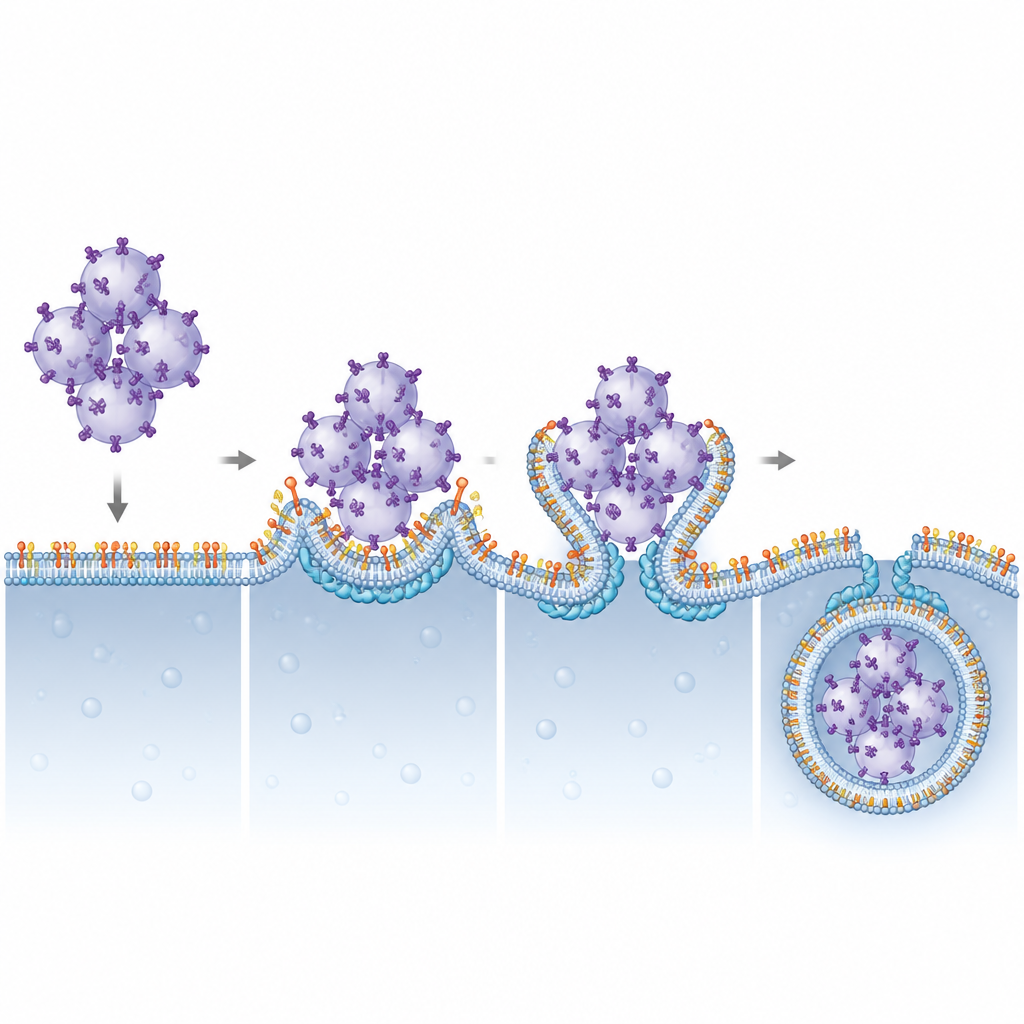
To visualize what went wrong when Pikfyve was blocked, the team used advanced fluorescence and electron microscopy. Under normal conditions, activated STING travels to recycling endosomes, gets packaged into clusters of small vesicles, and these clusters are then swallowed by lysosomes. When Pikfyve was inhibited, lysosomes expanded in size but failed to engulf these STING-rich vesicle clusters. Instead, hundreds of small STING-containing vesicles piled up just outside lysosomes, even though STING still carried the molecular tags that normally mark it for disposal. This showed that the problem lay not in recognizing STING as trash, but in the final step where lysosomes wrap and internalize the cargo.
A Lipid–Protein Pair That Drives Membrane Swallowing
The authors next asked which ESCRT components rely on PI(3,5)P2 to work at lysosomes. By systematically reducing various ESCRT subunits, they pinpointed CHMP4B, part of the ESCRT-III group, as crucial for STING breakdown. CHMP4B normally forms dynamic filaments that can constrict and cut membranes. Imaging showed that CHMP4B sits on lysosome membranes and that this localization disappears when Pikfyve or PI(3,5)P2 production is blocked. Computer simulations and biochemical tests revealed that a small cluster of positively charged amino acids on CHMP4B specifically recognizes and binds PI(3,5)P2. Mutating this cluster prevented CHMP4B from attaching to lysosomes, stopped it from binding PI(3,5)P2, and failed to rescue STING degradation or signaling shut-off in cells lacking normal CHMP4B.
Why This Matters for Immunity and Disease
This work outlines a clear structural and functional partnership between a rare membrane lipid, PI(3,5)P2, and the ESCRT-III protein CHMP4B in turning off the STING alarm. By anchoring CHMP4B on lysosomes, PI(3,5)P2 allows these organelles to bend their membranes around STING-loaded vesicle clusters and pinch them into their interior for destruction. When this system is disturbed, STING signaling persists, which could help explain inflammatory features in disorders linked to Pikfyve function and suggests that carefully tuning this lipid–protein axis might one day help boost anticancer immunity or rein in harmful inflammation.
Citation: Shoji, T., Shinojima, A., Kishimoto, T. et al. A PI(3,5)P2/CHMP4B axis on lysosomes is essential for microautophagic degradation of STING. Nat Commun 17, 4602 (2026). https://doi.org/10.1038/s41467-026-72828-4
Keywords: STING, lysosomal microautophagy, PI(3,5)P2, ESCRT-III, innate immunity