Clear Sky Science · es
Un eje PI(3,5)P2/CHMP4B en lisosomas es esencial para la degradación por microautofagia de STING
Cómo las células calman una alarma interna
Nuestras células disponen de un sistema de alarma interno que detecta ADN fuera de lugar, un signo de peligro asociado a infección viral o daño celular. Un interruptor clave llamado STING ayuda a convocar defensas antivirales e inflamatorias. Pero, como toda alarma, STING debe apagarse cuando la amenaza se ha resuelto, o puede impulsar inflamación crónica y enfermedad. Este estudio revela cómo componentes diminutos de membrana en los centros de reciclaje celular, los lisosomas, actúan en conjunto para engullir y desmantelar STING, restituyendo la calma celular.
La alarma de ADN de la célula y sus riesgos
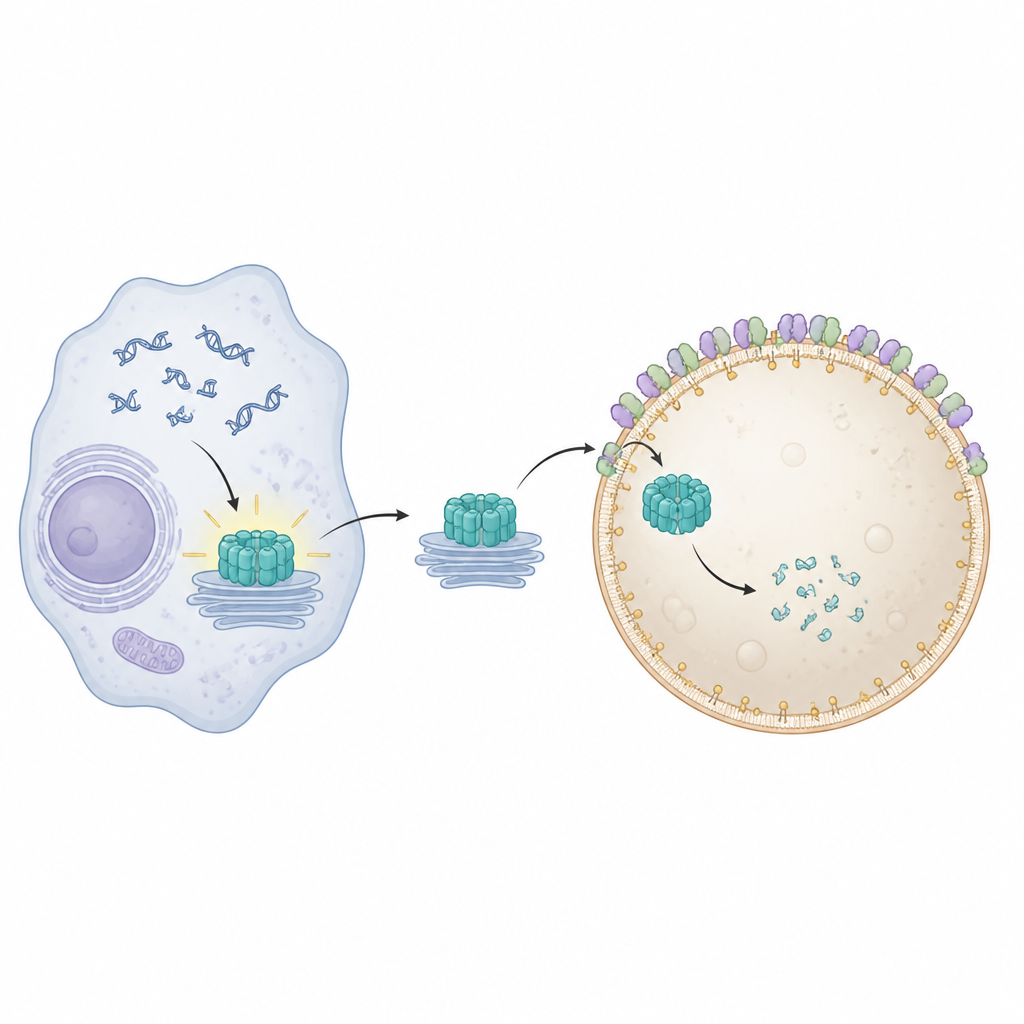
STING se localiza en membranas internas y se activa al detectar ADN en un lugar indebido dentro de la célula. Una vez activado, se traslada desde su ubicación habitual cerca del núcleo a través de varias estaciones de membrana y desencadena la producción de moléculas antivirales e inflamatorias. Para evitar daños duraderos, las células deben eliminar STING usado con rapidez. Trabajos anteriores mostraron que proteínas de remodelado de membrana especiales, conocidas colectivamente como ESCRT, ayudan a que los lisosomas pinzen y engullan carga directamente desde su superficie en un proceso llamado microautofagia. Sin embargo, no se había aclarado cómo STING queda físicamente envuelto y capturado por los lisosomas.
Encontrando un interruptor molecular de apagado
Los investigadores primero desarrollaron una prueba sensible que rastrea la velocidad a la que las células degradan STING al fusionarlo con enzimas productores de luz. Luego aplicaron una batería de inhibidores de quinasas, fármacos que bloquean distintas enzimas de señalización, para ver cuáles ralentizaban la degradación de STING. Varios compuestos destacaron, en particular los que inhiben una enzima llamada Pikfyve. Pikfyve contribuye a sintetizar un lípido señalizador raro, PI(3,5)P2, en compartimentos tardíos celulares, incluidos los lisosomas. Inhibir Pikfyve redujo PI(3,5)P2, impidió la degradación eficiente de STING y provocó que las señales activadas de STING persistieran, con expresión prolongada de genes inflamatorios tanto en células inmunitarias de ratón como humanas.
Viendo cómo STING queda fuera del reciclador celular
Para visualizar qué fallaba cuando se bloqueó Pikfyve, el equipo empleó microscopía de fluorescencia avanzada y microscopía electrónica. En condiciones normales, STING activado viaja a endosomas de reciclaje, se empaqueta en cúmulos de pequeñas vesículas, y estos cúmulos son luego engullidos por lisosomas. Cuando se inhibió Pikfyve, los lisosomas se agrandaron pero no lograron engullir esos cúmulos de vesículas ricas en STING. En su lugar, cientos de pequeñas vesículas que contienen STING se acumularon justo fuera de los lisosomas, aun cuando STING conservaba las etiquetas moleculares que normalmente lo marcan para eliminación. Esto indicó que el problema no residía en reconocer a STING como residuo, sino en el paso final en el que los lisosomas envuelven e internalizan la carga.
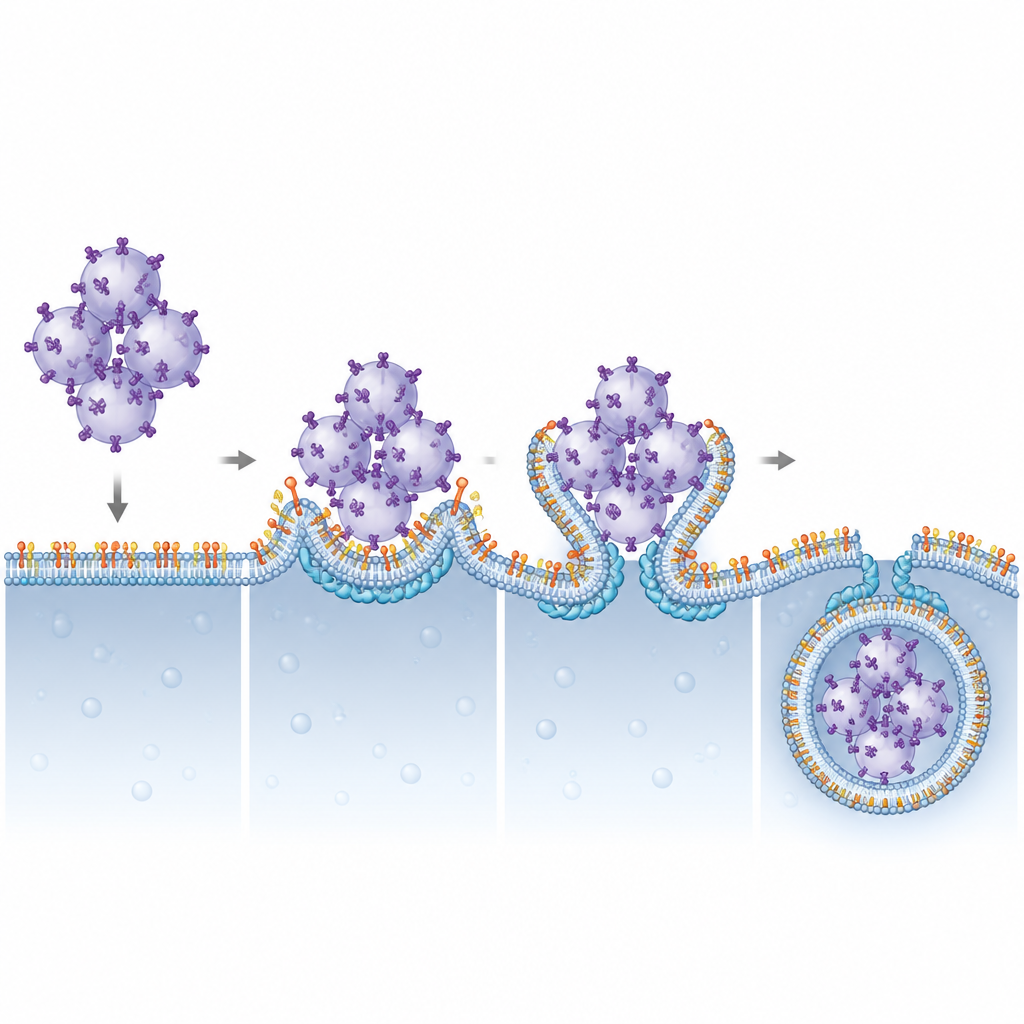
Una pareja lípido–proteína que impulsa el engullimiento de membrana
Los autores investigaron a continuación qué componentes de ESCRT dependen de PI(3,5)P2 para actuar en lisosomas. Al reducir sistemáticamente diversas subunidades de ESCRT, identificaron a CHMP4B, parte del grupo ESCRT-III, como crucial para la degradación de STING. CHMP4B normalmente forma filamentos dinámicos capaces de contraer y cortar membranas. Las imágenes mostraron que CHMP4B se localiza en las membranas lisosomales y que esta localización desaparece cuando se bloquea Pikfyve o la producción de PI(3,5)P2. Simulaciones por ordenador y ensayos bioquímicos revelaron que un pequeño cúmulo de aminoácidos cargados positivamente en CHMP4B reconoce y se une específicamente a PI(3,5)P2. Mutar este cúmulo impidió que CHMP4B se anclara en los lisosomas, evitó su unión a PI(3,5)P2 y no logró restaurar la degradación de STING ni el apagado de la señalización en células carentes de CHMP4B normal.
Por qué esto importa para la inmunidad y la enfermedad
Este trabajo perfila una asociación estructural y funcional clara entre un lípido de membrana poco frecuente, PI(3,5)P2, y la proteína ESCRT-III CHMP4B en el apagado de la alarma STING. Al anclar CHMP4B en los lisosomas, PI(3,5)P2 permite que estos orgánulos curven sus membranas alrededor de cúmulos de vesículas cargadas con STING y los pinzen hacia su interior para su destrucción. Cuando este sistema se altera, la señalización de STING persiste, lo que podría ayudar a explicar rasgos inflamatorios en trastornos relacionados con la función de Pikfyve y sugiere que ajustar con precisión este eje lípido–proteína podría algún día ayudar a potenciar la inmunidad anticancerígena o controlar la inflamación dañina.
Cita: Shoji, T., Shinojima, A., Kishimoto, T. et al. A PI(3,5)P2/CHMP4B axis on lysosomes is essential for microautophagic degradation of STING. Nat Commun 17, 4602 (2026). https://doi.org/10.1038/s41467-026-72828-4
Palabras clave: STING, microautofagia lisosomal, PI(3,5)P2, ESCRT-III, inmunidad innata