Clear Sky Science · fr
Un axe PI(3,5)P2/CHMP4B sur les lysosomes est essentiel pour la dégradation microautophagique de STING
Comment les cellules calment une alarme interne
Nos cellules disposent d’un système d’alarme interne qui détecte l’ADN errant, un signe de danger indiquant une infection virale ou des dommages cellulaires. Un interrupteur clé de cette alarme, appelé STING, contribue à mobiliser des défenses antivirales et inflammatoires. Mais, comme toute alarme, STING doit être désactivé une fois la menace neutralisée, sinon il peut provoquer une inflammation chronique et des maladies. Cette étude révèle comment de minuscules composants membranaires sur des centres de recyclage cellulaires appelés lysosomes coopèrent pour engloutir et démanteler STING, restaurant ainsi le calme cellulaire.
L’alarme à ADN de la cellule et ses risques
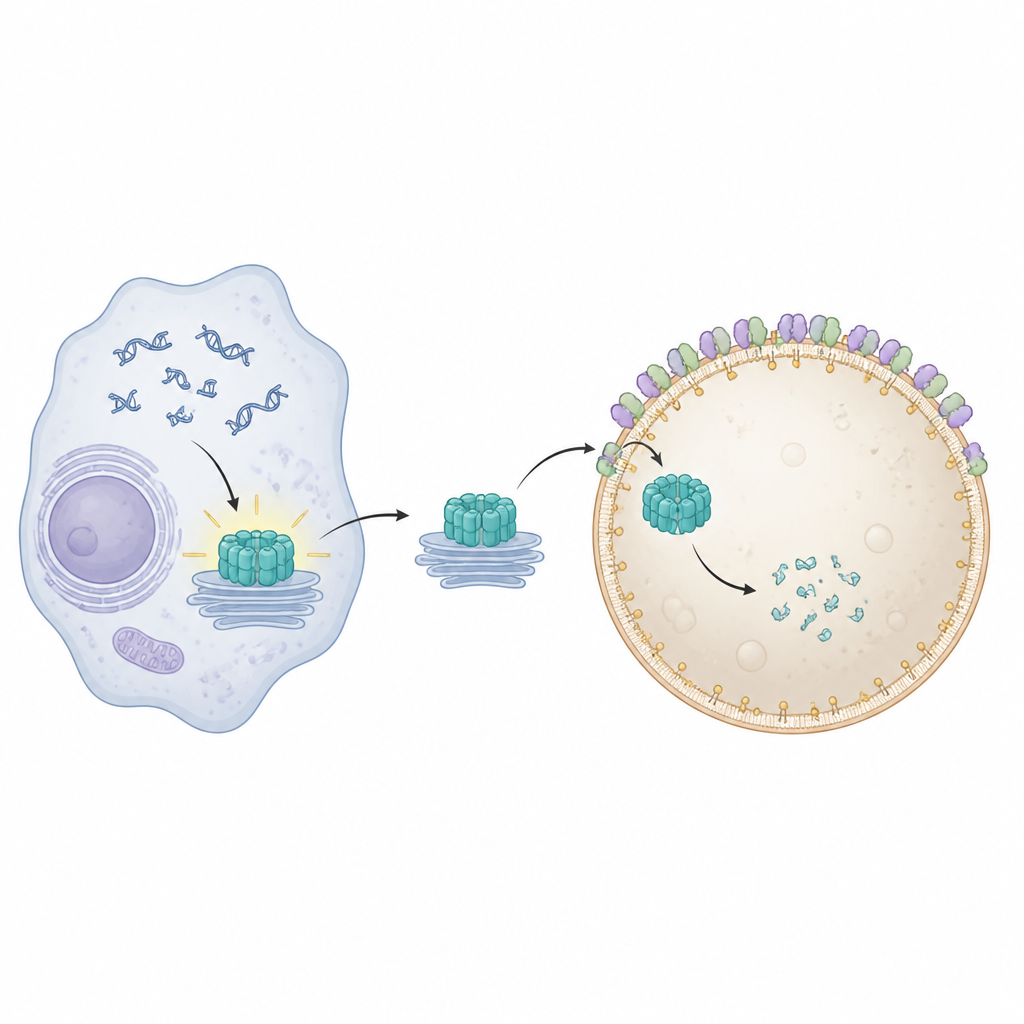
STING se trouve sur des membranes internes et s’active lorsqu’il détecte de l’ADN au mauvais endroit à l’intérieur d’une cellule. Une fois activé, il migre depuis son emplacement habituel près du noyau à travers plusieurs stations membranaires et déclenche la production de molécules antivirales et inflammatoires. Pour éviter des dommages durables, les cellules doivent éliminer rapidement le STING utilisé. Des travaux antérieurs ont montré que des protéines spécialisées de remodelage membranaire, connues collectivement sous le nom d’ESCRT, aident les lysosomes à pincer et engloutir directement des cargaisons à partir de leur surface dans un processus appelé microautophagie. Cependant, la façon dont STING est physiquement enroulé et internalisé par les lysosomes restait floue.
Trouver un interrupteur moléculaire d’arrêt
Les chercheurs ont d’abord mis au point un test sensible qui suit la vitesse de dégradation de STING en le fusionnant à des enzymes productrices de lumière. Ils ont ensuite appliqué une série d’inhibiteurs de kinases, des médicaments qui bloquent de nombreuses protéines de signalisation, pour identifier lesquels ralentissaient la dégradation de STING. Plusieurs composés se sont distingués, en particulier ceux qui bloquent une enzyme nommée Pikfyve. Pikfyve participe à la synthèse d’un lipide de signalisation rare, PI(3,5)P2, sur des compartiments cellulaires tardifs, y compris les lysosomes. L’inhibition de Pikfyve réduisait le PI(3,5)P2, empêchait la dégradation efficace de STING et faisait persister les signaux STING activés, entraînant une expression prolongée de gènes inflammatoires aussi bien dans des cellules immunitaires de souris que d’humain.
Observer STING coincé à l’extérieur du centre de recyclage de la cellule
Pour visualiser ce qui dysfonctionnait lorsque Pikfyve était bloqué, l’équipe a utilisé des microscopes de fluorescence et électroniques avancés. En conditions normales, STING activé migre vers des endosomes de recyclage, est empaqueté en grappes de petites vésicules, et ces grappes sont ensuite avalées par les lysosomes. Quand Pikfyve était inhibé, les lysosomes augmentaient de taille mais ne parvenaient pas à engloutir ces grappes vésiculaires riches en STING. À la place, des centaines de petites vésicules contenant STING s’accumulaient juste à l’extérieur des lysosomes, bien que STING porte encore les marques moléculaires qui le ciblent normalement pour l’élimination. Cela montrait que le problème ne venait pas de la reconnaissance du STING comme déchet, mais de l’étape finale où les lysosomes enveloppent et internalisent la cargaison.
Une paire lipide–protéine qui propulse l’engloutissement membranaire
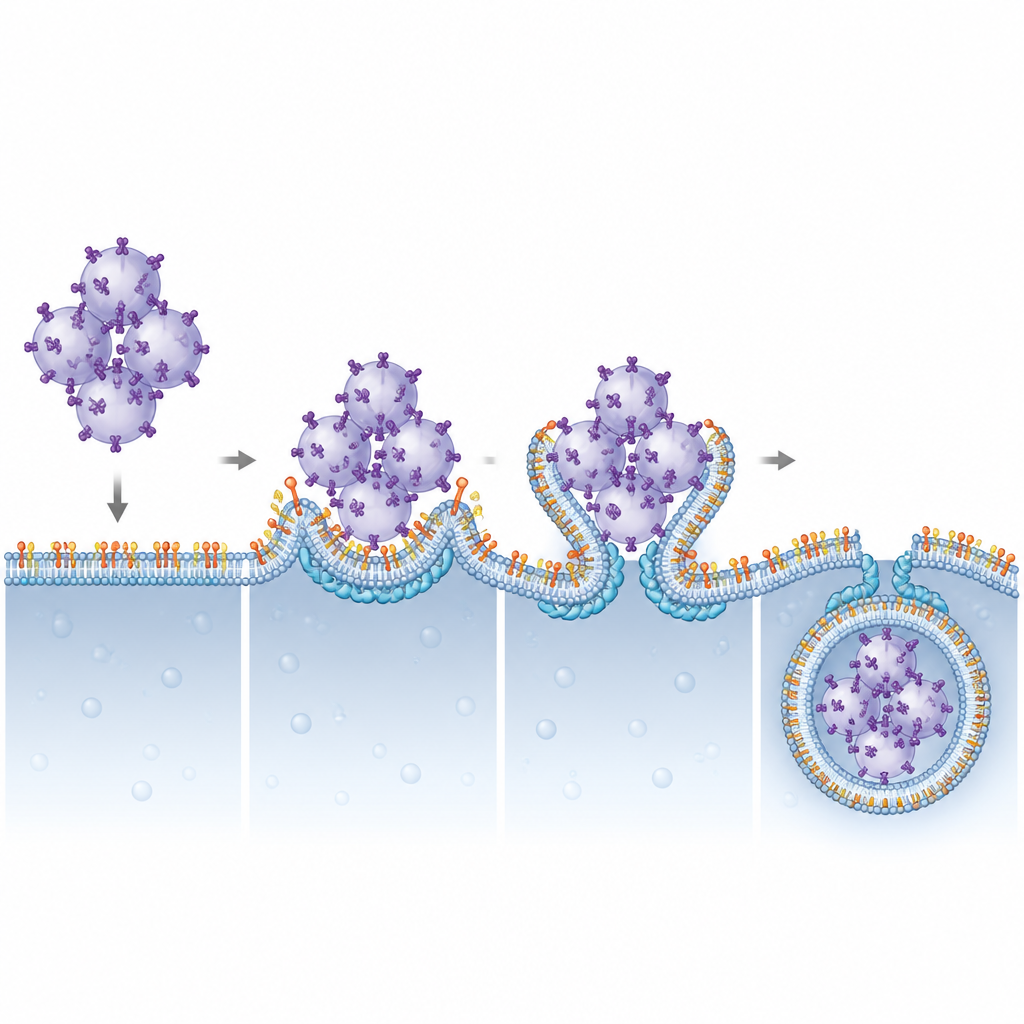
Les auteurs se sont ensuite demandés quels composants de l’ESCRT dépendent du PI(3,5)P2 pour agir sur les lysosomes. En réduisant systématiquement diverses sous-unités de l’ESCRT, ils ont identifié CHMP4B, membre du groupe ESCRT-III, comme crucial pour la dégradation de STING. CHMP4B forme normalement des filaments dynamiques capables de constricter et de découper les membranes. L’imagerie montrait que CHMP4B se localise sur les membranes lysosomales et que cette localisation disparaît lorsque Pikfyve ou la production de PI(3,5)P2 est bloquée. Des simulations informatiques et des tests biochimiques ont révélé qu’un petit groupe d’acides aminés chargés positivement sur CHMP4B reconnaît et se lie spécifiquement au PI(3,5)P2. La mutation de ce groupe empêchait CHMP4B de s’attacher aux lysosomes, le privait de liaison au PI(3,5)P2 et n’a pas permis de restaurer la dégradation de STING ni l’arrêt de la signalisation dans des cellules dépourvues de CHMP4B normal.
Pourquoi cela compte pour l’immunité et la maladie
Ce travail décrit un partenariat structurel et fonctionnel clair entre un lipide membranaire rare, le PI(3,5)P2, et la protéine ESCRT-III CHMP4B pour éteindre l’alarme STING. En ancrant CHMP4B sur les lysosomes, le PI(3,5)P2 permet à ces organites de courber leurs membranes autour de grappes de vésicules chargées de STING et de les pincer dans leur intérieur pour les détruire. Quand ce système est perturbé, la signalisation STING persiste, ce qui pourrait expliquer certaines manifestations inflammatoires associées à des troubles liés à la fonction de Pikfyve, et suggère que l’ajustement fin de cet axe lipide–protéine pourrait un jour aider à renforcer l’immunité anticancéreuse ou à limiter des inflammations nuisibles.
Citation: Shoji, T., Shinojima, A., Kishimoto, T. et al. A PI(3,5)P2/CHMP4B axis on lysosomes is essential for microautophagic degradation of STING. Nat Commun 17, 4602 (2026). https://doi.org/10.1038/s41467-026-72828-4
Mots-clés: STING, microautophagie lysosomale, PI(3,5)P2, ESCRT-III, immunité innée