Clear Sky Science · ru
Крио-ЭМ структура Photosystem I Chlamydomonas reinhardtii в комплексе с цитохромом c6
Как зелёные водоросли поддерживают поток солнечного света
Каждый зелёный лист и клетка водоросли зависят от невидимой эстафеты электронов, превращающей свет в пригодную для использования энергию. В этом исследовании рассматривается одно крошечное «передаваемое эстафетное» действие: как небольшой переносчик — белок цитохром c6 — доставляет электроны к гигантскому светопоглощающему аппарату, известному как Фотосистема I, у зелёных водорослей. Замораживая эти молекулы в момент взаимодействия и визуализируя их с почти атомным разрешением, авторы показывают, как происходит это ключевое столкновение и как оно связано с эволюционной историей фотосинтеза.
Тщательно выстроенная цепочка энергетических шагов
В кислородпроизводящем фотосинтезе энергия течёт через цепь больших белковых комплексов, встроенных во внутренние мембраны. Фотосистема II использует свет, чтобы расщепить воду и передать электроны на промежуточную станцию — комплекс цитохром b6f. Оттуда маленькие растворимые переносчики — либо меди-содержащий пластоцианин, либо железосодержащий цитохром c6 — перевозят электроны через водную среду к Фотосистеме I. Когда Фотосистема I получает эти электроны, она помогает генерировать электрические и химические градиенты, которые в конечном счёте питают синтез АТФ — универсальной энергетической валюты клетки.
Древний партнёр встречает современную машину
С эволюционной точки зрения предполагается, что самые ранние версии Фотосистемы I в основном работали с цитохромом c6. Со временем многие организмы, особенно растения, перешли на использование пластоцианина, частично потому, что он содержит медь, а не железо, которого может не хватать. Зелёные водоросли, такие как Chlamydomonas reinhardtii, занимают среднюю позицию в этой истории: они могут использовать оба переносчика и меняют предпочтение в зависимости от доступности металлов. Понимание того, как именно цитохром c6 прикрепляется к Фотосистеме I у этих водорослей, даёт окно в то, как древние и современные стратегии доставки электронов сосуществуют и как они могли эволюционировать.
Замораживание момента контакта
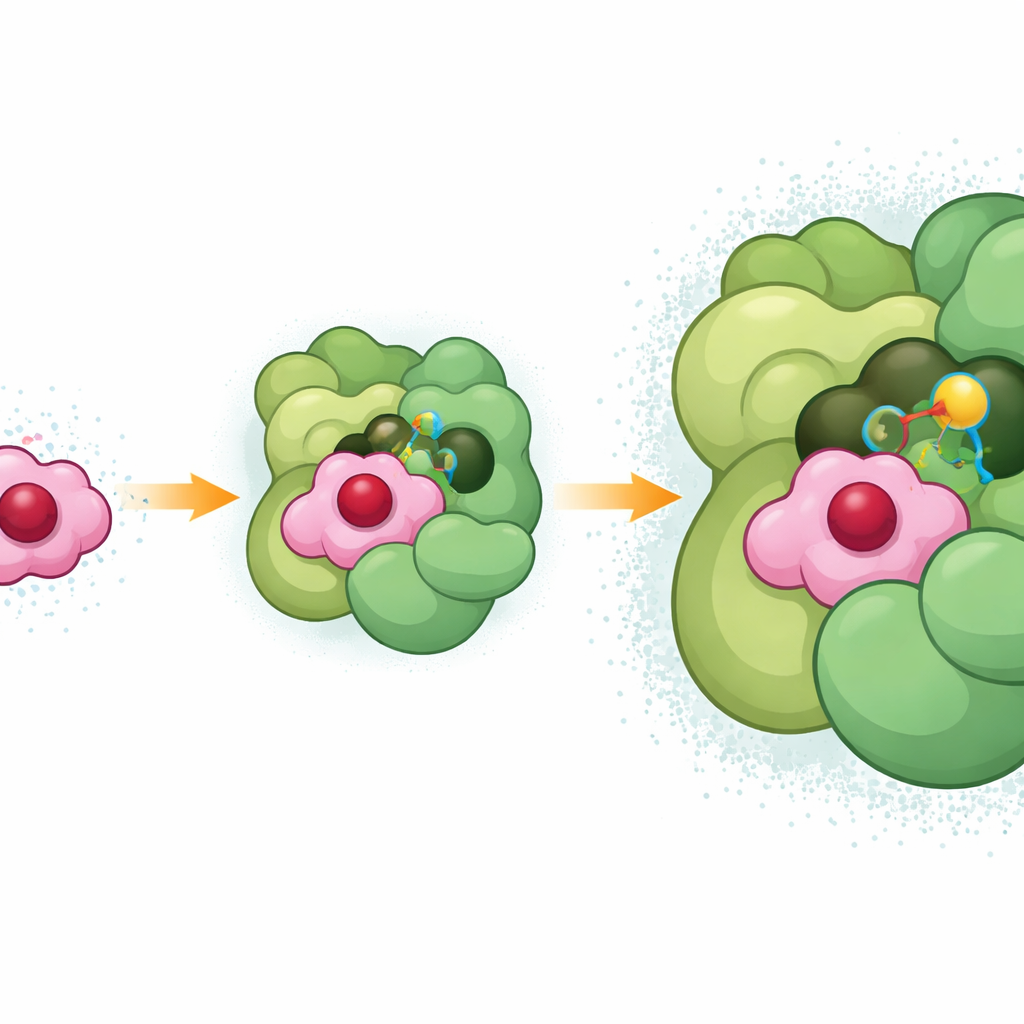
Исследователи создали стабильные комплексы между цитохромом c6 и Фотосистемой I, используя кросссвязывающий реагент короткого радиуса действия, который соединяет два партнёра только когда они естественно находятся рядом. Затем они применили высокоразрешающую крио-электронную микроскопию для реконструкции трёхмерной структуры этого замороженного взаимодействия с разрешением около двух ангстремов — достаточно тонким, чтобы разместить отдельные боковые цепи аминокислот, молекулы пигментов и даже многие молекулы воды. Структура показывает цитохром c6, устроенный в неглубокой впадине, образованной там, где встречаются два основных субъединицы Фотосистемы I, а дополнительная поверхностная спираль другой субъединицы (PsaF) нависает словно маленькая рука, помогая зафиксировать его на месте.
Точная подгонка для быстрого потока электронов
В этой впадине гем цитохрома c6 — часть, которая фактически переносит электрон — располагается всего примерно в одиннадцати ангстремах от особой пары хлорофиллов в Фотосистеме I, принимающей электрон. Интерфейс поддерживается плотной сетью взаимодействий: отрицательно заряженные участки цитохрома c6 притягивают положительные области на PsaF, тогда как нейтральные и ароматические боковые цепи создают плотные, «маслянистые» контакты глубже в канавке. Конкретная аргинин-остаток в цитохроме c6 (R66), давно известный как важный у бактерий, устанавливает стопку с соседними ароматическими кольцами как в цитохроме c6, так и в Фотосистеме I, формируя трёхслойное взаимодействие, которое, по-видимому, помогает стабилизировать позицию докинга. Такая плотная упаковка также отталкивает воду от прямого пути между гемом и хлорофиллом, вероятно снижая сопротивление потоку электронов и объясняя, почему перенос электронов происходит всего за несколько миллионных долей секунды.
Проверка ключевых точек контакта
Чтобы выяснить, какие части цитохрома c6 наиболее важны, команда ввела целевые замены кислотных и основных остатков, предположительно контактирующих с Фотосистемой I. Когда они нейтрализовали определённые отрицательные заряды вблизи «руки» PsaF или заменили ключевой аргинин, скорость перерегенерации Фотосистемы I заметно замедлялась, а кросссвязывание между двумя белками ослабевало. Некоторые комбинации изменений вызывали неожидаемое поведение, намекая на то, что цитохром c6 и Фотосистема I иногда могут собираться в альтернативные, менее эффективные конформации. Вместе эти тесты подтверждают, что и электростатический захват на основе PsaF, и аргинин-центрированное сайтинговое взаимодействие критически важны для быстрого и надёжного переноса электронов.
Что это значит для солнечных машин жизни
Работа предоставляет детальную структурную схему того, как цитохром c6 передаёт электроны Фотосистеме I у зелёных водорослей, сочетая черты, наблюдаемые у бактерий, с характеристиками более продвинутых растений. Она показывает, как небольшой переносчик и крупный мембранный комплекс коэволюционировали тонко настроенный интерфейс, уравновешивающий сильное связывание, быстрый перенос электронов и быстрое высвобождение для следующего цикла. Уточняя этот древний шаг в фотосинтетической эстафете, исследование помогает объяснить, как организмы оптимизировали преобразование света в энергию в ходе эволюции, и даёт подсказки для будущих попыток проектировать или имитировать природные солнечные энергетические системы.
Цитирование: Ogawa, Y., Mahapatra, G.P., Milrad, Y. et al. Cryo-EM structure of Chlamydomonas reinhardtii Photosystem I complexed with cytochrome c6. Nat Commun 17, 3031 (2026). https://doi.org/10.1038/s41467-026-70944-9
Ключевые слова: фотосинтез, Фотосистема I, цитохром c6, перенос электронов, крио-ЭМ