Clear Sky Science · pt
Interações competitivas moldam a dinâmica e a computação das redes cerebrais de mamíferos
Por que isso importa para entender a mente
O cérebro costuma ser imaginado como uma teia harmoniosa de atividade, com regiões trabalhando juntas para pensar, sentir e agir. Mas cérebros reais também estão cheios de empurra-e-puxa: algumas áreas se reforçam mutuamente, enquanto outras puxam em direções opostas. Este estudo investiga como essa guerra silenciosa molda o funcionamento dos cérebros de mamíferos — incluindo humanos, macacos e camundongos — e mostra que a competição entre regiões não é um defeito, mas uma característica vital que torna a atividade cerebral mais realista, mais individualizada e mais capaz de computação complexa.
Como a fiação do cérebro orienta cooperação e rivalidade
Os autores partem de um enigma simples: o diagrama estrutural do cérebro, ou conectoma, informa sobretudo onde as fibras passam, não se essas conexões agem como ajudantes amigáveis ou rivais opostos. A maioria dos modelos cerebrais em larga escala assume silenciosamente que, se uma região conectada fica mais ativa, tende a aumentar a atividade de seus parceiros também. Porém a biologia é repleta de interações competitivas — de neurônios inibitórios em circuitos locais até feedbacks de equilíbrio em ecossistemas e redes sociais. A equipe, portanto, constrói um modelo de todo o cérebro que pode atribuir a cada conexão de longa distância uma influência positiva (cooperativa) ou negativa (competitiva) e pergunta qual mistura melhor reproduz os padrões reais de atividade medidos por ressonância funcional em humanos, macacos e camundongos.
Construindo cérebros virtuais a partir da anatomia real
Para isso, os pesquisadores usam um instrumento matemático padrão na neurociência de redes: um conjunto de osciladores não lineares próximos a um ponto de inflexão, um por região cerebral. Esses osciladores produzem naturalmente sinais oscilantes e flutuantes semelhantes às ondas lentas observadas em fMRI, nem completamente ruidosos nem perfeitamente regulares. Eles acoplam os osciladores de acordo com as conexões estruturais de cada espécie — a partir de difusão por ressonância magnética em humanos, dados de macacos que mesclam rastreamento de tratos e imageamento, e rastreamento puro de tratos em camundongos — e então ajustam iterativamente a força e o sinal desses links existentes para que a atividade simulada corresponda aos padrões observados de coativação ao longo do tempo. O resultado é uma matriz de “conectividade generativa”: uma versão funcionalizada do diagrama de fiação que captura como a estrutura deve ser ponderada para dar origem à dinâmica do cérebro vivo.
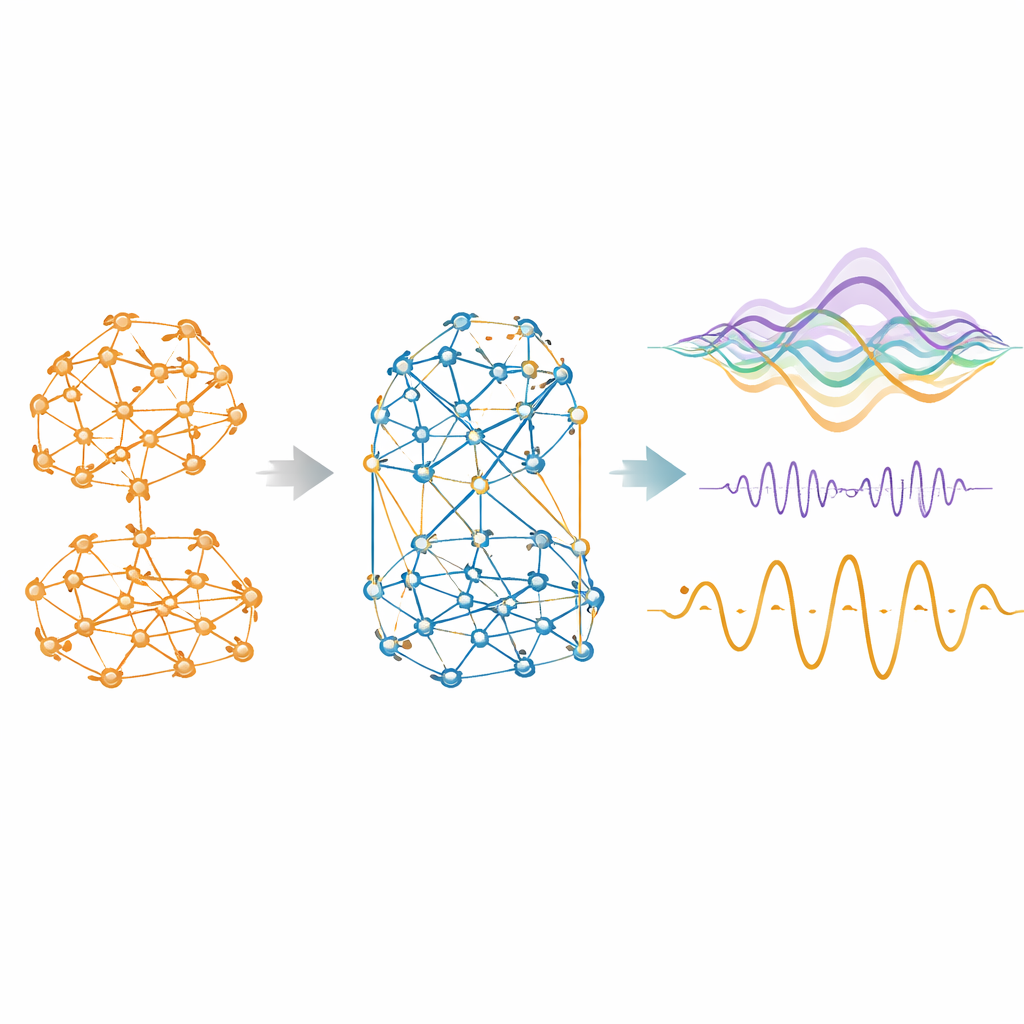
A competição emerge como um ingrediente necessário
Quando o modelo pode usar influências negativas além das positivas, ele o faz com entusiasmo. Entre as espécies, cerca de um quarto a dois quintos das conexões efetivas tornam-se competitivas. Esses links negativos não são distribuídos ao acaso: tendem a ser mais fracos, de longo alcance e menos agrupados do que os cooperativos, formando uma teia difusa que se entrecruza com comunidades positivas mais modulares. Removê-los degrada rapidamente a capacidade do modelo de corresponder aos dados reais. Com a competição incluída, a similaridade entre a conectividade funcional simulada e a empírica salta dramaticamente, alcançando correlações de até 0,95 em camundongos e mais do que dobrando em humanos em comparação com um modelo apenas positivo. O modelo aprimorado também produz níveis realistas de anticorrelação entre regiões, reproduzindo os padrões de ‘balança’ amplamente observados em exames cerebrais.
A competição se alinha a diferenças biológicas profundas
A equipe então investiga se a localização dos links competitivos reflete uma organização biológica mais profunda. Eles comparam onde as conexões negativas do modelo aparecem com mapas de várias características corticais, incluindo tipos celulares, expressão gênica, distribuição de receptores, microestrutura e mielinização. Em humanos, macacos e camundongos, os links competitivos preferencialmente conectam regiões que se situam em extremos opostos desses gradientes biológicos — por exemplo, áreas ricas em um tipo de interneurônio inibitório versus áreas ricas em outro, ou zonas sensoriais altamente mielinizadas versus córtex de associação transmodais. Em efeito, a grande disputa em larga escala é mais forte entre territórios corticais que são construídos e sintonizados de maneira muito diferente, sugerindo que a competição macroscópica está ancorada em contrastes moleculares e celulares conservados.
Dinâmica mais rica e cérebros mais individuais
Incluir competição faz mais do que melhorar fotos estáticas da conectividade. Quando os modelos ajustados são executados para gerar atividade cerebral sintética, a versão cooperativa–competitiva mostra um comportamento temporal mais realista. Ela evita uma sincronia global irrealisticamente alta, em vez disso vagando por um regime equilibrado que alterna integração e segregação, uma propriedade conhecida como metastabilidade. Também exibe hierarquia mais forte, onde algumas regiões estão melhor posicionadas para desencadear cascatas amplas de atividade ou para enviar mais informação do que recebem. Medidas de informação sinérgica — a ideia de que pares de regiões juntos carregam mais poder preditivo do que cada uma isoladamente — aumentam em direção a níveis empíricos. Crucialmente, essas melhorias não foram explicitamente alvo durante o ajuste; elas emergem naturalmente uma vez permitidas as interações competitivas.
Da atividade realista à computação e cognição
Como nenhum cérebro é exatamente igual a outro, um modelo convincente deve ser ao mesmo tempo fiel e pessoal. Os modelos cooperativo–competitivos provaram ser mais “identificáveis”: eles se ajustam melhor à conectividade de cada indivíduo do que à de outros e sofrem uma queda maior na precisão quando atribuídos ao sujeito errado, em humanos, macacos e camundongos. Para humanos, os autores também testaram quão bem os padrões instantâneos de atividade do modelo correspondiam a grandes bases de mapas cerebrais vinculadas a funções mentais específicas, como atenção ou memória. Modelos com competição produziram padrões espontâneos que se assemelhavam mais a esses circuitos cognitivos canônicos, implicando dinâmicas mais “semelhantes à mente” em repouso. Finalmente, quando as redes derivadas do modelo foram usadas como fiação em sistemas artificiais de “reservoir computing” executando uma tarefa de memória, aquelas com links competitivos mostraram maior capacidade computacional, retenção melhor das entradas passadas.
O que isso significa para entender cérebros e construir modelos
Em termos claros, o estudo mostra que o cérebro de mamíferos funciona melhor num ponto de equilíbrio entre cooperação e competição. Vínculos positivos fortes e locais unem regiões próximas em módulos especializados, enquanto vínculos negativos mais fracos e de longo alcance opõem diferentes sistemas entre si, estruturando o fluxo de informação pelo córtex. Essa arquitetura não apenas reproduz as peculiaridades finas de cérebros individuais, mas também gera naturalmente dinâmicas que são diversas, hierárquicas e poderosas do ponto de vista computacional. Para modelagem cerebral futura — seja para entender a consciência, simular doenças ou projetar computadores neuromórficos — a mensagem é clara: deixar de fora a competição em larga escala é deixar de fora um princípio central de como cérebros reais computam.
Citação: Luppi, A.I., Sanz Perl, Y., Vohryzek, J. et al. Competitive interactions shape mammalian brain network dynamics and computation. Nat Neurosci 29, 915–933 (2026). https://doi.org/10.1038/s41593-026-02205-3
Palavras-chave: redes cerebrais, competição neural, modelagem do conectoma, conectividade funcional, computação neuromórfica