Clear Sky Science · pl
Interakcje konkurencyjne kształtują dynamikę i przetwarzanie w sieciach mózgów ssaków
Dlaczego to ma znaczenie dla rozumienia umysłu
Mózgi często wyobrażamy sobie jako harmonijne sieci aktywności, gdzie obszary współpracują, by myśleć, czuć i działać. Jednak prawdziwe mózgi są pełne pociągnięć i przeciągnięć: niektóre regiony wzajemnie się wzmacniają, podczas gdy inne ciągną w przeciwnych kierunkach. W pracy tej bada się, jak ten ukryty przeciąg wpływa na sposób działania mózgów ssaków — w tym ludzi, makaków i myszy — i pokazuje, że konkurencja między obszarami mózgu nie jest błędem, lecz istotną cechą, która sprawia, że aktywność mózgu jest bardziej realistyczna, bardziej indywidualna i zdolna do złożonych obliczeń.
Jak okablowanie mózgu kieruje współpracą i rywalizacją
Autorzy zaczynają od prostego problemu: strukturalny schemat połączeń mózgu, czyli connectom, w większości mówi nam, gdzie biegną włókna, a nie czy te połączenia działają jak przyjazni wspomagacze, czy jak przeciwnicy. Większość modeli wielkoskalowych zakłada bez słowa, że jeśli jeden połączony region staje się bardziej aktywny, skłania to jego partnerów ku wzrostowi aktywności. Tymczasem biologia jest pełna interakcji konkurencyjnych — od neuronów hamujących w lokalnych obwodach po równoważące sprzężenia zwrotne w ekosystemach i sieciach społecznych. Zespół zatem buduje model całego mózgu, który może przypisać każdemu połączeniu dalekiego zasięgu wpływ dodatni (współpraca) lub ujemny (konkurencja), i pyta, która mieszanka najlepiej odtwarza rzeczywiste wzorce aktywności zmierzone za pomocą fMRI u ludzi, makaków i myszy.
Budowanie wirtualnych mózgów z realnej anatomii
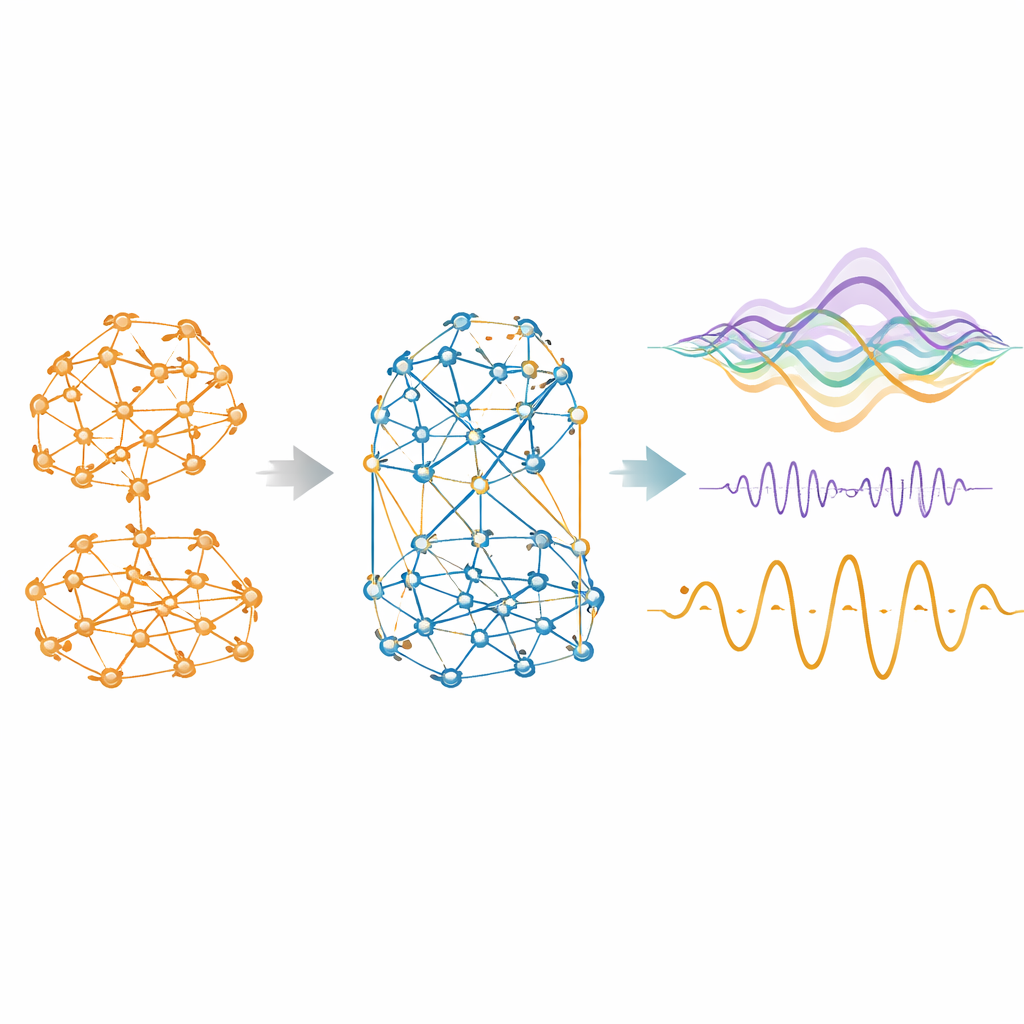
Aby to osiągnąć, badacze używają standardowego narzędzia matematycznego w neurobiologii sieciowej: zestawu nieliniowych oscylatorów ustawionych blisko punktu przełomowego, po jednym na region mózgu. Te oscylatory naturalnie generują fluktuujące, rytmiczne sygnały podobne do wolnych fal obserwowanych w fMRI, nie całkowicie losowe ani idealnie regularne. Sprzęgają oscylatory zgodnie ze strukturą połączeń dla każdego gatunku — z ludzkiego MRI dyfuzyjnego, danych makaka łączących śledzenie włókien i obrazowanie oraz czystego śledzenia tras u myszy — a następnie iteracyjnie dostosowują siłę i znak istniejących połączeń, tak by symulowana aktywność odpowiadała obserwowanym wzorcom współaktywacji w czasie. Wynikiem jest macierz „generatywnej łączności”: sfunkcjonalizowana wersja schematu okablowania, która oddaje, jak struktura musi być ważona, by rodziła dynamikę żywego mózgu.
Konkurencja wysuwa się jako niezbędny składnik
Gdy model może używać wpływów ujemnych obok dodatnich, robi to chętnie. Wśród gatunków mniej więcej jedna czwarta do dwóch piątych efektywnych połączeń staje się konkurencyjna. Te ujemne łącza nie są rozsypywane losowo: mają tendencję do bycia słabszymi, długozasięgowymi i mniej ściśle skupionymi niż te współpracujące, tworząc rozproszoną sieć, która przecina bardziej modułowe, dodatnie społeczności. Usunięcie ich szybko pogarsza zgodność modelu z rzeczywistymi danymi. Po uwzględnieniu konkurencji podobieństwo między symulowaną a empiryczną łącznością funkcjonalną skacze dramatycznie, osiągając korelacje nawet do 0,95 u myszy i bardziej niż podwajając się u ludzi w porównaniu z modelem tylko z dodatnimi wpływami. Ulepszony model generuje też realistyczne poziomy antykorelacji między regionami, odpowiadające powszechnie obserwowanym wzorcom „huśtawki” w skanach mózgu.
Konkurencja pokrywa się z głębszymi różnicami biologicznymi
Zespół następnie bada, czy rozmieszczenie konkurencyjnych łączy odzwierciedla głębszą organizację biologiczną. Porównują miejsca, gdzie pojawiają się ujemne połączenia modelowe, z mapami wielu cech korowych, w tym typów komórek, ekspresji genów, rozmieszczenia receptorów, mikrostuktury i mielinizacji. Wśród ludzi, makaków i myszy połączenia konkurencyjne preferencyjnie łączą regiony znajdujące się na przeciwległych końcach tych gradientów biologicznych — na przykład obszary bogate w jeden rodzaj interneuronu hamującego kontra obszary bogate w inny, lub silnie zmielinizowane strefy sensoryczne kontra kora asocjacyjna transmodalna. W efekcie wielkoskalowe przeciąganie liny jest najsilniejsze między terytoriami korowymi zbudowanymi i dostrojonymi bardzo odmiennie, co sugeruje, że makroskalowa konkurencja jest zakotwiczona w zachowanych kontrastach molekularnych i komórkowych.
Bogatsza dynamika i bardziej indywidualne mózgi
Uwzględnienie konkurencji robi więcej niż poprawia statyczne migawki łączności. Gdy dopasowane modele są uruchamiane do przodu, by generować syntetyczną aktywność mózgu, wersja kooperacyjno‑konkurencyjna wykazuje bardziej realistyczne zachowanie czasowe. Unika nierealistycznie wysokiej synchronii globalnej, zamiast tego oscylując w zrównoważonym reżimie, który przeplata integrację i segregację — właściwość znaną jako metastabilność. Pokazuje też silniejszą hierarchię, gdzie niektóre regiony są lepiej ustawione, by zapoczątkować rozległe kaskady aktywności lub wysyłać więcej informacji, niż otrzymują. Miary informacji synergicznej — idea, że para regionów razem niesie więcej mocy predykcyjnej niż każdy z osobna — rosną w kierunku poziomów empirycznych. Co istotne, te ulepszenia nie były explicite celowane podczas dopasowywania; wyłaniają się naturalnie, gdy pozwoli się na interakcje konkurencyjne.
Od realistycznej aktywności do obliczeń i poznania
Ponieważ nie ma dwóch identycznych mózgów, przekonujący model musi być wierny i osobisty. Modele kooperacyjno‑konkurencyjne okazały się bardziej „odciskowalne”: lepiej pasowały do łączności konkretnej osoby niż do innych i traciły więcej na dokładności, gdy były użyte dla niewłaściwego osobnika — u ludzi, makaków i myszy. U ludzi autorzy sprawdzili też, jak dobrze chwilowe wzory aktywności modelu odpowiadają dużym bazom map mózgu powiązanych z określonymi funkcjami umysłowymi, takimi jak uwaga czy pamięć. Modele z konkurencją generowały spontaniczne wzory bliższe tym kanonicznym obwodom poznawczym, co sugeruje bardziej „umysłopodobną” dynamikę w spoczynku. Wreszcie, gdy sieci wyprowadzone z modelu zostały użyte jako okablowanie w sztucznych systemach „reservoir computing” rozwiązujących zadanie pamięciowe, te z połączeniami konkurencyjnymi wykazały wyższą pojemność obliczeniową, lepiej utrzymując ślady poprzednich wejść.
Co to znaczy dla rozumienia mózgów i budowy modeli
Mówiąc prosto, badanie pokazuje, że mózg ssaków działa najlepiej na ostrzu noża między współpracą a konkurencją. Silne, lokalne więzi dodatnie łączą pobliskie regiony w wyspecjalizowane moduły, podczas gdy słabsze, długozasięgowe więzi ujemne przeciwstawiają różne systemy, strukturując przepływ informacji przez korę. Ta architektura nie tylko odtwarza drobne osobnicze niuanse mózgów, ale też naturalnie daje dynamikę, która jest zróżnicowana, hierarchiczna i potężna obliczeniowo. Dla przyszłego modelowania mózgu — czy to w celu zrozumienia świadomości, symulacji chorób, czy projektowania komputerów neuromorficznych — przekaz jest jasny: pominięcie wielkoskalowej konkurencji oznacza pominięcie zasadniczej zasady działania prawdziwych mózgów.
Cytowanie: Luppi, A.I., Sanz Perl, Y., Vohryzek, J. et al. Competitive interactions shape mammalian brain network dynamics and computation. Nat Neurosci 29, 915–933 (2026). https://doi.org/10.1038/s41593-026-02205-3
Słowa kluczowe: sieci mózgowe, konkurencja neuronalna, modelowanie connectomu, łączność funkcjonalna, obliczenia neuromorficzne