Clear Sky Science · fr
Les interactions compétitives façonnent la dynamique et le calcul des réseaux cérébraux des mammifères
Pourquoi c’est important pour comprendre l’esprit
On imagine souvent le cerveau comme une toile harmonieuse d’activités, où les régions coopèrent pour penser, ressentir et agir. Mais les cerveaux réels sont aussi pleins de tensions : certaines zones se renforcent mutuellement, tandis que d’autres tirent dans des directions opposées. Cette étude examine comment cette lutte invisible façonne le fonctionnement des cerveaux de mammifères — humains, macaques et souris — et montre que la compétition entre régions n’est pas un défaut mais une caractéristique essentielle qui rend l’activité cérébrale plus réaliste, plus individuelle et plus apte à des calculs complexes.
Comment l’anatomie guide coopération et rivalité
Les auteurs partent d’un constat simple : le schéma structurel du cerveau, ou connectome, indique principalement où passent les fibres, mais pas si ces connexions agissent comme des aides amicales ou des rivales opposées. La plupart des modèles cérébraux à grande échelle supposent tacitement que si une région connectée devient plus active, elle tend aussi à augmenter l’activité de ses partenaires. Pourtant, la biologie regorge d’interactions compétitives — des neurones inhibiteurs dans les circuits locaux aux rétroactions équilibrantes dans les écosystèmes et les réseaux sociaux. L’équipe construit donc un modèle cérébral complet qui peut attribuer à chaque connexion longue distance une influence positive (coopérative) ou négative (compétitive) et cherche quelle combinaison reproduit le mieux les motifs d’activité mesurés par IRMf chez l’humain, le macaque et la souris.
Construire des cerveaux virtuels à partir de l’anatomie réelle
Pour cela, les chercheurs utilisent un outil mathématique standard en neurosciences des réseaux : un ensemble d’oscillateurs non linéaires placés près d’un point de basculement, un par région cérébrale. Ces oscillateurs produisent naturellement des signaux fluctuants et rythmiques similaires aux ondes lentes observées en IRMf, ni complètement bruités ni parfaitement réguliers. Ils couplent les oscillateurs selon les connexions structurelles propres à chaque espèce — issues de l’IRM de diffusion chez l’humain, de données macaques mêlant traçage de fibres et imagerie, et de traçage pur chez la souris — puis ajustent itérativement la force et le signe de ces liaisons existantes pour que l’activité simulée corresponde aux motifs d’activation conjointe observés au fil du temps. Le résultat est une matrice de « connectivité générative » : une version fonctionnalisée du schéma de câblage qui capture comment il faut pondérer la structure pour engendrer la dynamique du cerveau vivant.
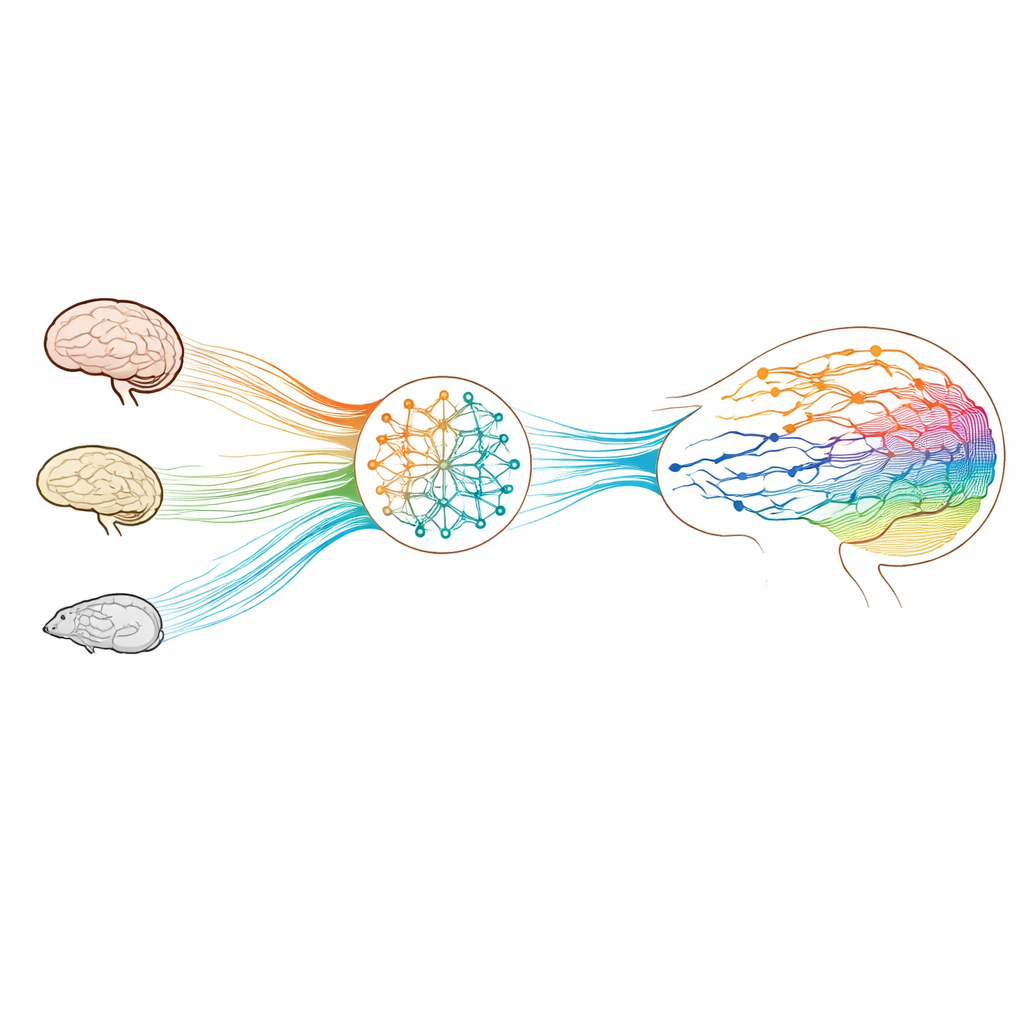
La compétition émerge comme un ingrédient nécessaire
Lorsque le modèle peut utiliser des influences négatives aussi bien que positives, il s’en sert avec enthousiasme. Selon les espèces, environ un quart à deux cinquièmes des connexions effectives deviennent compétitives. Ces liens négatifs ne sont pas répartis au hasard : ils ont tendance à être plus faibles, de plus longue portée et moins fortement regroupés que les liens coopératifs, formant une toile diffuse qui traverse les communautés positives plus modulaires. Les supprimer dégrade rapidement l’adéquation du modèle aux données réelles. Avec la compétition incluse, la similarité entre la connectivité fonctionnelle simulée et empirique augmente fortement, atteignant des corrélations allant jusqu’à 0,95 chez la souris et plus que doublant chez l’humain par rapport à un modèle uniquement positif. Le modèle amélioré produit aussi des niveaux réalistes d’anticorrélation entre régions, reproduisant les motifs en « bascule » longtemps observés dans les scans cérébraux.
La compétition correspond à des différences biologiques profondes
L’équipe se demande ensuite si la localisation des liens compétitifs reflète une organisation biologique plus fondamentale. Ils comparent la position des connexions négatives du modèle avec des cartes de nombreuses caractéristiques corticales, incluant types cellulaires, expression génique, distribution des récepteurs, microstructure et myélinisation. Chez l’humain, le macaque et la souris, les liens compétitifs relient préférentiellement des régions situées aux extrémités opposées de ces gradients biologiques — par exemple des zones riches en un type d’interneurone inhibiteur contre des zones riches en un autre, ou des zones sensorielles fortement myélinisées contre le cortex associatif transmodal. En pratique, la lutte à grande échelle est la plus forte entre des territoires corticaux construits et réglés très différemment, ce qui suggère que la compétition macroscopique est ancrée dans des contrastes moléculaires et cellulaires conservés.
Des dynamiques plus riches et des cerveaux plus individuels
Inclure la compétition fait plus qu’améliorer des clichés statiques de la connectivité. Quand les modèles ajustés sont exécutés pour générer une activité cérébrale synthétique, la version coopérative–compétitive montre un comportement temporel plus réaliste. Elle évite une synchronie globale irréaliste et oscille plutôt dans un régime équilibré alternant intégration et ségrégation, une propriété connue sous le nom de métastabilité. Elle affiche aussi une hiérarchie plus marquée, où certaines régions sont mieux placées pour déclencher des cascades d’activité étendues ou pour envoyer plus d’information qu’elles n’en reçoivent. Les mesures d’information synergique — l’idée que deux régions ensemble portent plus de pouvoir prédictif que chacune séparément — s’élèvent vers des niveaux empiriques. De manière cruciale, ces améliorations n’étaient pas explicitement ciblées pendant l’ajustement ; elles émergent naturellement dès que les interactions compétitives sont autorisées.
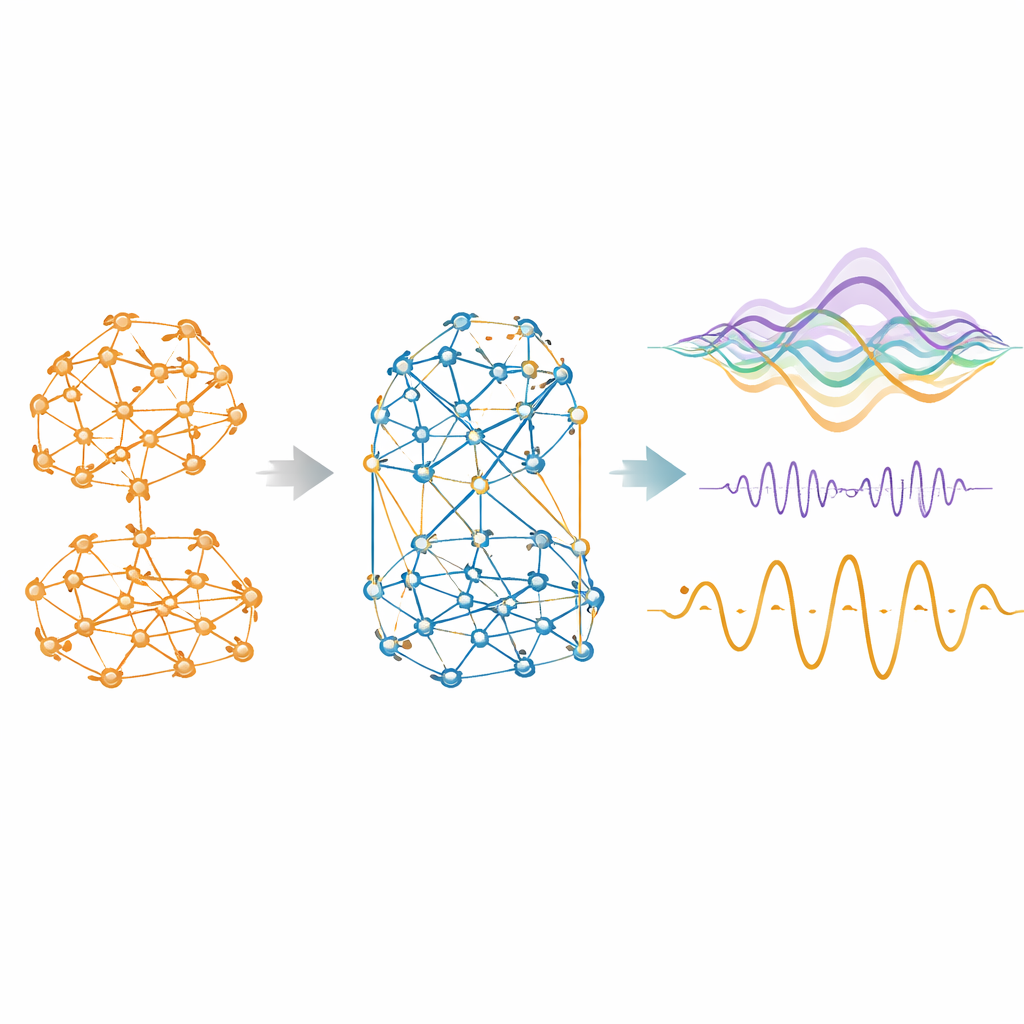
De l’activité réaliste au calcul et à la cognition
Parce que deux cerveaux ne sont jamais identiques, un modèle convaincant doit être à la fois fidèle et personnel. Les modèles coopératifs–compétitifs se sont révélés plus « empreintables » : ils correspondent mieux à la connectivité de chaque individu qu’à celle des autres et subissent une plus forte perte de précision lorsqu’ils sont appliqués au mauvais sujet, chez l’humain comme chez le macaque et la souris. Pour l’humain, les auteurs ont aussi testé dans quelle mesure les motifs d’activité instantanée du modèle ressemblaient à de grandes bases de données de cartes cérébrales associées à des fonctions mentales spécifiques, comme l’attention ou la mémoire. Les modèles avec compétition produisent des motifs spontanés qui ressemblent davantage à ces circuits cognitifs canoniques, impliquant des dynamiques de repos plus « semblables à l’esprit ». Enfin, lorsque les réseaux dérivés du modèle ont servi de câblage dans des systèmes artificiels de « reservoir computing » effectuant une tâche de mémoire, ceux incorporant des liens compétitifs ont montré une capacité de calcul supérieure, retenant mieux les entrées passées.
Ce que cela signifie pour comprendre le cerveau et construire des modèles
En termes simples, l’étude montre que le cerveau des mammifères fonctionne au mieux à la lisière entre coopération et compétition. De forts liens positifs locaux rassemblent les régions proches en modules spécialisés, tandis que des liens négatifs plus faibles et de longue portée opposent différents systèmes, structurant le flux d’information à travers le cortex. Cette architecture non seulement reproduit les particularités fines des cerveaux individuels, mais génère aussi naturellement des dynamiques diverses, hiérarchiques et puissantes sur le plan computationnel. Pour la modélisation future du cerveau — qu’il s’agisse de comprendre la conscience, simuler la maladie ou concevoir des ordinateurs neuromorphiques — le message est clair : omettre la compétition à grande échelle revient à laisser de côté un principe central du calcul cérébral réel.
Citation: Luppi, A.I., Sanz Perl, Y., Vohryzek, J. et al. Competitive interactions shape mammalian brain network dynamics and computation. Nat Neurosci 29, 915–933 (2026). https://doi.org/10.1038/s41593-026-02205-3
Mots-clés: réseaux cérébraux, compétition neuronale, modélisation du connectome, connectivité fonctionnelle, informatique neuromorphique