Clear Sky Science · it
Le interazioni competitive plasmano la dinamica e il calcolo delle reti cerebrali dei mammiferi
Perché è importante per comprendere la mente
I cervelli vengono spesso immaginati come reti armoniche di attività, con aree che collaborano per pensare, sentire e agire. Ma i cervelli reali sono anche pieni di spinte contrapposte: alcune aree si potenziano a vicenda, mentre altre tirano in direzioni opposte. Questo studio indaga come quella lotta nascosta plasmi il funzionamento effettivo dei cervelli dei mammiferi — inclusi esseri umani, macachi e topi — e mostra che la competizione tra regioni cerebrali non è un difetto ma una caratteristica vitale che rende l’attività cerebrale più realistica, più individuale e più capace di calcoli complessi.
Come l’anatomia guida cooperazione e rivalità
Gli autori partono da un puzzle semplice: il diagramma strutturale delle connessioni cerebrali, o connettoma, ci dice soprattutto dove corrono le fibre, non se quelle connessioni agiscono come alleati o come avversari. La maggior parte dei modelli cerebrali su larga scala assume implicitamente che se una regione connessa aumenta la propria attività, tenderà a far aumentare anche i partner. Eppure la biologia è piena di interazioni competitive — dai neuroni inibitori nei circuiti locali al feedback di bilanciamento negli ecosistemi e nelle reti sociali. Il gruppo costruisce quindi un modello cerebrale globale che può assegnare a ogni connessione a lunga distanza un’influenza positiva (cooperativa) o negativa (competitiva) e verifica quale miscela riproduce meglio i pattern di attività misurati con la risonanza magnetica funzionale in umani, macachi e topi.
Costruire cervelli virtuali a partire dall’anatomia reale
Per farlo, i ricercatori usano un classico strumento matematico nelle neuroscienze delle reti: un insieme di oscillatori non lineari posti vicino a un punto critico, uno per ciascuna regione cerebrale. Questi oscillatori producono naturalmente segnali fluttuanti e ritmici simili alle onde lente osservate con la fMRI, né completamente casuali né perfettamente regolari. Collegano gli oscillatori secondo le connessioni strutturali di ciascuna specie — dalla dMRI umana, ai dati del macaco che combinano tracciamento di tratti e imaging, fino al puro tracciamento di tratti nei topi — e poi aggiustano iterativamente la forza e il segno di quei collegamenti esistenti in modo che l’attività simulata corrisponda ai pattern osservati di co-attivazione nel tempo. Il risultato è una matrice di “connettività generativa”: una versione funzionalizzata del diagramma delle connessioni che cattura come la struttura deve essere pesata per dare origine alla dinamica del cervello vivo.
La competizione emerge come ingrediente necessario
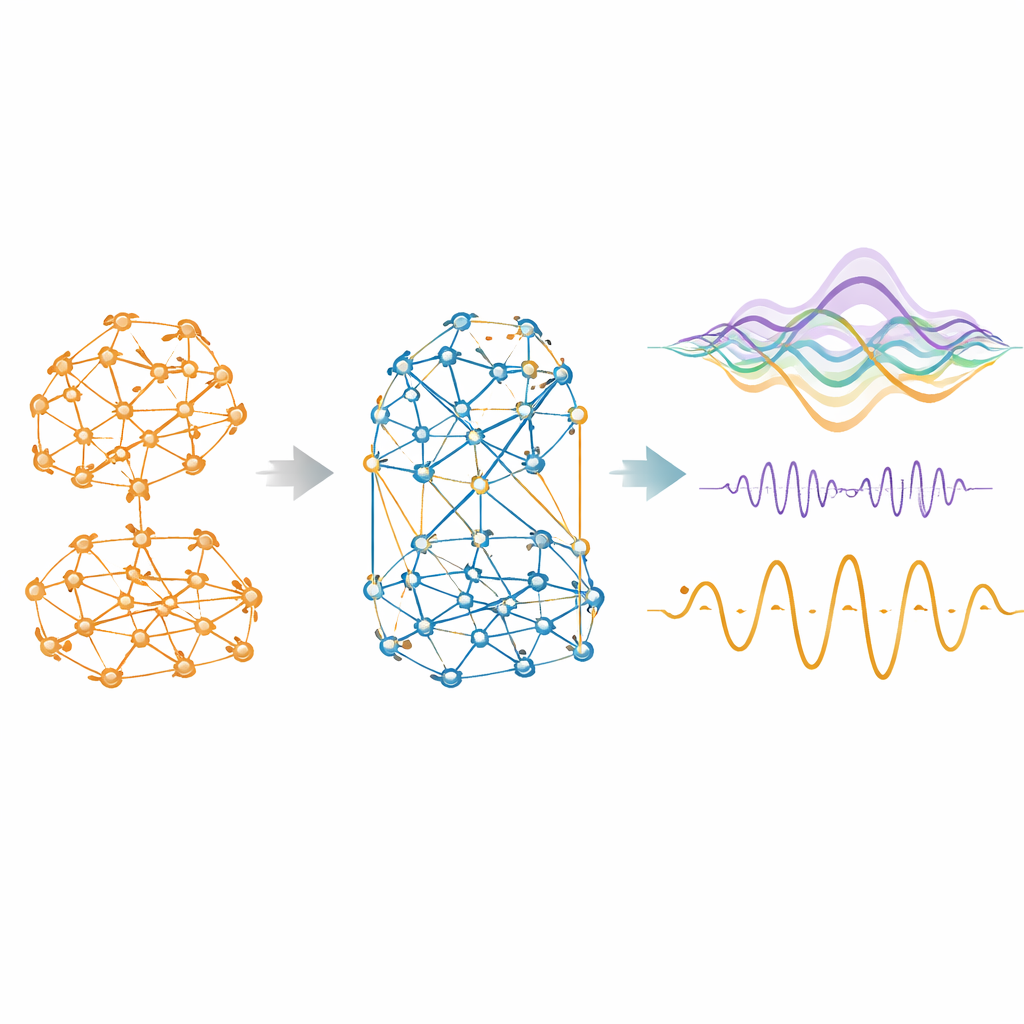
Quando al modello si permette di usare influenze negative oltre che positive, lo fa con decisione. Tra le specie, all’incirca un quarto fino a due quinti delle connessioni efficaci diventano competitive. Questi legami negativi non sono distribuiti a caso: tendono a essere più deboli, a lungo raggio e meno strettamente raggruppati rispetto a quelli cooperativi, formando una rete diffusa che attraversa le comunità positive più modulari. Rimuoverli degrada rapidamente la capacità del modello di riprodurre i dati reali. Con la competizione inclusa, la somiglianza tra connettività funzionale simulata ed empirica aumenta drasticamente, raggiungendo correlazioni fino a 0,95 nei topi e più che raddoppiando negli esseri umani rispetto a un modello solo positivo. Il modello migliorato produce anche livelli realistici di anticorrelazione tra regioni, rispecchiando i diffusi schemi a “bilanciere” osservati a lungo nelle scansioni cerebrali.
La competizione si allinea a profonde differenze biologiche
Il gruppo chiede poi se la collocazione dei legami competitivi rifletta una organizzazione biologica più profonda. Confrontano dove cadono le connessioni negative del modello con mappe di molte caratteristiche corticali, inclusi tipi cellulari, espressione genica, distribuzione dei recettori, microstruttura e mielinizzazione. In umani, macachi e topi, i collegamenti competitivi tendono a unire preferenzialmente regioni poste agli estremi opposti di questi gradienti biologici — per esempio, aree ricche di un tipo di interneurone inibitorio rispetto ad aree ricche di un altro, o zone sensoriali fortemente mielinizzate rispetto alla corteccia associativa transmodale. Di fatto, la lotta su larga scala è più intensa tra territori corticali costruiti e sintonizzati in modo molto diverso, suggerendo che la competizione macroscopica sia ancorata a contrasti molecolari e cellulari conservati.
Dinamiche più ricche e cervelli più individuali
Includere la competizione fa più che migliorare istantanee statiche della connettività. Quando i modelli adattati vengono fatti evolvere per generare attività cerebrale sintetica, la versione cooperativa–competitiva mostra un comportamento temporale più realistico. Evita sincronie globali irrealisticamente alte, muovendosi invece attraverso un regime bilanciato che alterna integrazione e segregazione, una proprietà nota come metastabilità. Mostra anche una gerarchia più marcata, dove alcune regioni sono meglio posizionate per innescare ampie cascate di attività o per inviare più informazione di quanta ne ricevano. Le misure di informazione sinergica — l’idea che coppie di regioni insieme contengano più potere predittivo di ciascuna separatamente — si avvicinano ai livelli empirici. Crucialmente, questi miglioramenti non sono stati esplicitamente bersagliati durante l’adattamento; emergono naturalmente una volta che si consentono interazioni competitive.
Dall’attività realistica al calcolo e alla cognizione
Poiché non esistono due cervelli identici, un modello convincente deve essere al tempo stesso fedele e personale. I modelli cooperativo–competitivi si sono dimostrati più “fingerprintabili”: si adattano meglio alla connettività di ciascun individuo rispetto a quella di altri e subiscono un calo di accuratezza maggiore quando abbinati all’individuo sbagliato, sia negli umani sia in macachi e topi. Per gli umani, gli autori hanno anche testato quanto bene i pattern di attività istantanei del modello corrispondessero a grandi banche dati di mappe cerebrali legate a funzioni mentali specifiche, come attenzione o memoria. I modelli con competizione hanno prodotto schemi spontanei che somigliano più da vicino a questi circuiti cognitivi canonici, implicando dinamiche a riposo più “simili alla mente”. Infine, quando le reti derivate dal modello sono state usate come cablaggio in sistemi artificiali di “reservoir computing” impegnati in un compito di memoria, quelle con collegamenti competitivi hanno mostrato una maggiore capacità computazionale, trattenendo meglio gli input passati.
Cosa significa per comprendere i cervelli e costruire modelli
In parole semplici, lo studio mostra che il cervello dei mammiferi funziona al meglio su un filo sottile tra cooperazione e competizione. Forti legami positivi locali legano regioni vicine in moduli specializzati, mentre legami negativi più deboli e a lungo raggio mettono in competizione sistemi diversi, strutturando il flusso d’informazione attraverso la corteccia. Questa architettura non solo riproduce le sottigliezze individuali dei cervelli, ma genera naturalmente dinamiche diverse, gerarchiche e potenti dal punto di vista computazionale. Per la modellizzazione cerebrale futura — sia per capire la coscienza, simulare le malattie o progettare computer neuromorfici — il messaggio è chiaro: escludere la competizione su larga scala significa omettere un principio centrale di come i cervelli reali computano.
Citazione: Luppi, A.I., Sanz Perl, Y., Vohryzek, J. et al. Competitive interactions shape mammalian brain network dynamics and computation. Nat Neurosci 29, 915–933 (2026). https://doi.org/10.1038/s41593-026-02205-3
Parole chiave: reti cerebrali, concorrenza neurale, modellizzazione del connettoma, connettività funzionale, calcolo neuromorfico