Clear Sky Science · pt
Fosforilação de SF3B1 por CDK11 orquestra ativação do spliceossomo via recrutamento do complexo RES dependente de SNIP1
Como as Células Editam Suas Mensagens Genéticas
Cada célula deve editar cuidadosamente as mensagens genéticas brutas antes de transformá-las em proteínas. Quando essa edição falha, pode contribuir para câncer e transtornos neurológicos. Este estudo revela como um pequeno grupo de proteínas dentro de nossas células atua como um interruptor temporal para garantir que as mensagens de RNA sejam processadas corretamente, e como uma única alteração herdada em uma dessas proteínas pode perturbar esse controle delicado.
A Máquina Celular de Edição
Nossos genes são primeiro copiados em longas fitas de RNA que contêm trechos úteis misturados com segmentos extras que precisam ser removidos. Uma enorme máquina celular chamada spliceossomo realiza esse corte e junção. Ela se monta em estágios, adicionando e liberando muitas peças proteicas enquanto avança de um estado inicial inativo até uma forma totalmente ativa. Uma peça central dessa máquina, uma proteína chamada SF3B1, é conhecida por ser quimicamente marcada e desmarcada durante o ciclo de edição, mas até agora os cientistas não compreendiam totalmente o que essas marcas fazem ou em que momento elas são mais importantes.
Encontrando um Passo de Edição Pausado
Para investigar essa questão, os pesquisadores usaram uma pequena molécula que bloqueia uma enzima chamada CDK11, que adiciona marcas químicas à SF3B1. Em células humanas tratadas com esse composto, eles isolaram partículas do spliceossomo ligadas ao DNA e mediram sua composição proteica. Descobriram um estado pausado até então desconhecido no ciclo de edição: um complexo que já tem alguns grupos de proteínas auxiliares no lugar enquanto outros ainda estão ausentes. Eles chamam essa forma arrestada de BOTS964. Nesse estado, um grupo auxiliar inicial já se juntou, mas um grupo posterior, normalmente necessário para a ativação completa, ainda não chegou, revelando um ponto de checagem específico no qual a atividade de CDK11 é necessária. 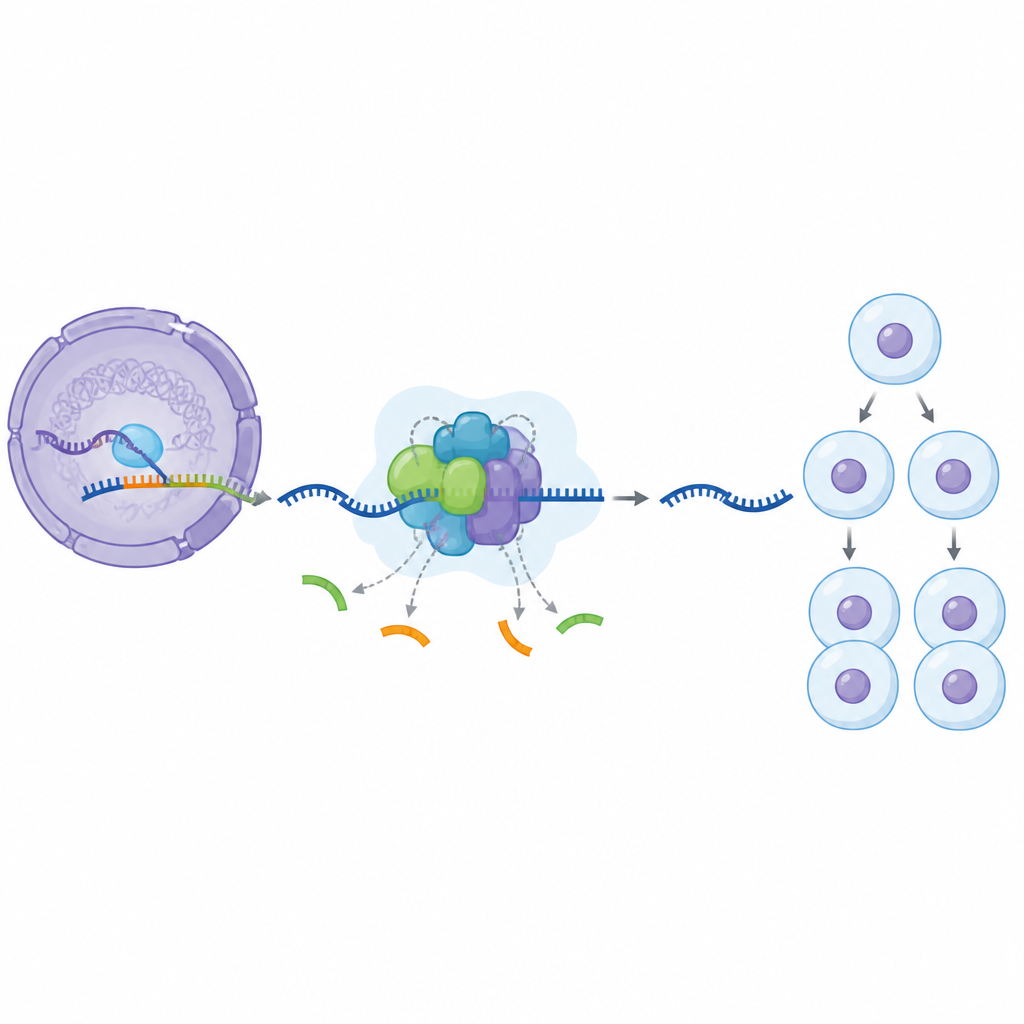
Como uma Marca em SF3B1 Molda o Sítio Ativo
A equipe então perguntou onde a versão fosforilada de SF3B1 realmente se posiciona no RNA. Usando uma técnica que entrelaça proteínas diretamente às bases de RNA que tocam, mapearam os pontos de contato da SF3B1 fosforilada dentro dos pequenos RNAs que formam o núcleo do spliceossomo. Eles descobriram que a SF3B1 marcada está enriquecida em um laço particular dentro do RNA U6, uma região que ajuda a moldar o núcleo catalítico onde ocorrem os cortes e junções. Quando a CDK11 foi bloqueada, esses contatos enfraqueceram, sugerindo que a adição de grupos fosfato à SF3B1 ajuda a estabilizar o centro de RNA dobrado necessário para um splicing preciso.
Uma Proteína Leitor Recruta um Complexo Auxiliar Chave
Em seguida, os cientistas buscaram proteínas que prefiram ligar-se à forma marcada de SF3B1. Eles identificaram SNIP1, parte de um conjunto de três proteínas auxiliares chamado complexo RES, conhecido por impedir que mensagens de RNA defeituosas escapem do núcleo. SNIP1 possui um domínio forkhead-associated, um bolso que reconhece marcas de fosfato específicas. Testes bioquímicos e modelagem estrutural mostraram que esse bolso interage com múltiplos sítios fosforilados dentro da cauda flexível da SF3B1. Essa interação ajuda a recrutar e ancorar o complexo RES completo ao spliceossomo justamente quando ele se torna cataliticamente ativo, garantindo uma progressão suave pela etapa de ativação.
Quando o Leitor Está Ausente ou Danificado
Para ver o que acontece quando o SNIP1 é removido subitamente, a equipe engenheirou células nas quais o SNIP1 pode ser degradado rapidamente. Em poucas horas de depleção, muitos genes apresentaram retenção generalizada de íntrons, indicando uma ampla falha no splicing de RNA. O padrão de defeitos correspondeu de perto ao observado quando a CDK11 é inibida, destacando que ambas as proteínas atuam juntas na mesma etapa da edição. Sem SNIP1, a maior parte do complexo RES não consegue se juntar ao spliceossomo, e a SF3B1 fica hiperfosforilada por CDK11, reforçando a ideia de que o recrutamento adequado de SNIP1 depende de uma SF3B1 corretamente fosforilada.
Ligação com um Transtorno Humano do Cérebro
Por fim, os pesquisadores examinaram mudanças sutis no bolso de SNIP1, incluindo uma mutação E366G previamente encontrada em pessoas de uma comunidade com um transtorno do neurodesenvolvimento. Proteínas SNIP1 mutantes ligaram-se menos à SF3B1 marcada, associaram-se com menos força ao spliceossomo ativo e foram menos capazes de restaurar o splicing e o crescimento celular quando o SNIP1 nativo foi removido. Outras mutações artificiais que enfraqueceram ainda mais essa interação causaram defeitos ainda mais graves. Em conjunto, esses resultados apoiam um modelo no qual a CDK11 primeiro marca a SF3B1, a SF3B1 marcada então recruta o SNIP1 e o complexo RES, e essa cadeia de eventos estabiliza o centro catalítico de RNA e mantém o splicing eficiente. Interromper qualquer elo dessa cadeia, incluindo mudanças associadas a doenças em SNIP1, pode comprometer o processamento de RNA e a função celular normal. 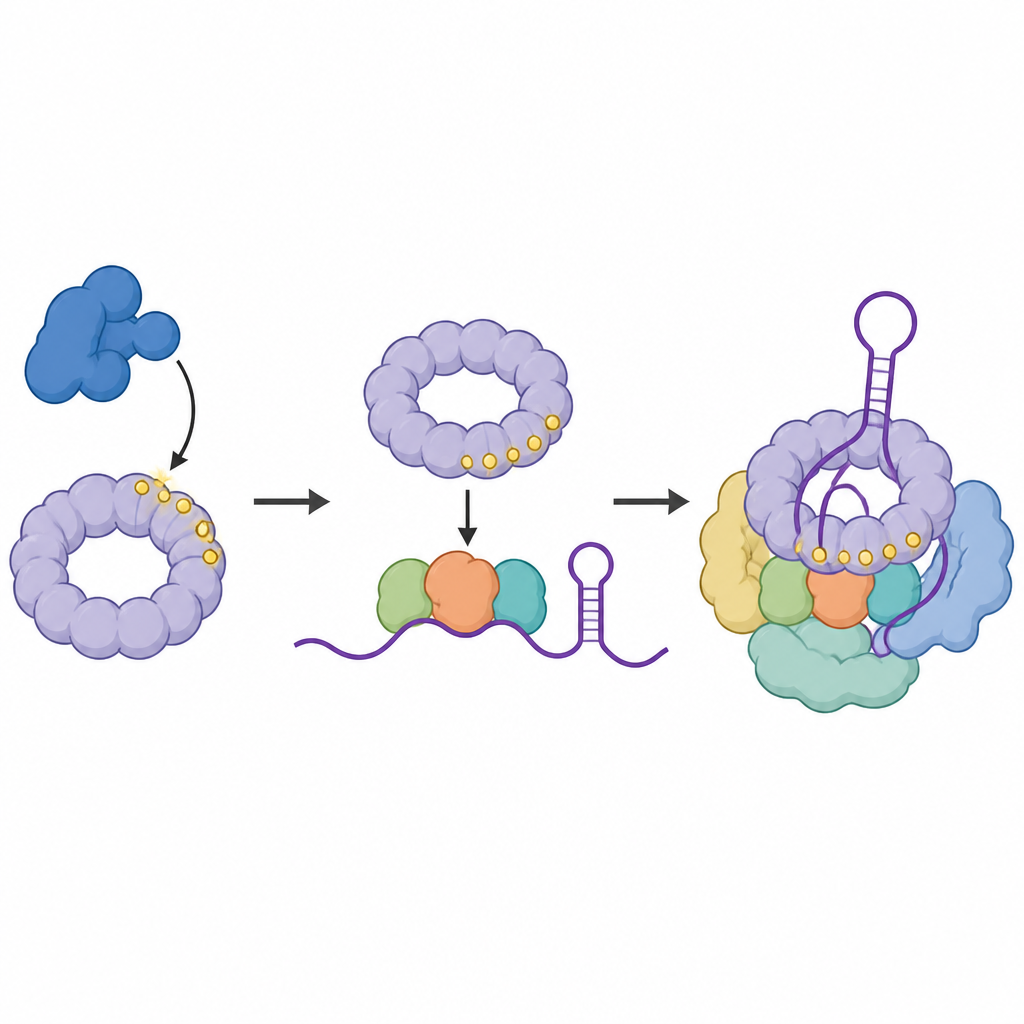
Citação: Gajdušková, P., Ruiz de Los Mozos, I., Hluchý, M. et al. Phosphorylation of SF3B1 by CDK11 orchestrates spliceosome activation via SNIP1-dependent RES complex recruitment. Nat Commun 17, 4577 (2026). https://doi.org/10.1038/s41467-026-71119-2
Palavras-chave: Splicing de RNA, spliceossomo, SF3B1, CDK11, SNIP1