Clear Sky Science · de
Phosphorylierung von SF3B1 durch CDK11 orchestriert die Aktivierung des Spleißosoms über SNIP1‑abhängige Rekrutierung des RES‑Komplexes
Wie Zellen ihre genetischen Botschaften bearbeiten
Jede Zelle muss rohe genetische Botschaften sorgfältig bearbeiten, bevor sie in Proteine übersetzt werden. Misslingt diese Bearbeitung, kann das zu Krebs und neurologischen Erkrankungen beitragen. Diese Studie deckt auf, wie eine kleine Gruppe von Proteinen in unseren Zellen als Zeitgeber zusammenarbeitet, um sicherzustellen, dass RNA‑Botschaften korrekt verarbeitet werden, und wie eine einzelne vererbte Veränderung in einem dieser Proteine diese feine Steuerung stören kann.
Die zelluläre Bearbeitungsmaschine
Unsere Gene werden zunächst in lange RNA‑Stränge kopiert, die nützliche Abschnitte mit zusätzlichen Segmenten mischen, die entfernt werden müssen. Eine riesige zelluläre Maschine, das Spleißosom, übernimmt dieses Schneiden und Verknüpfen. Es baut sich in Stufen auf und fügt viele Proteinanteile hinzu und setzt sie wieder frei, während es von einem frühen inaktiven Zustand in eine voll aktive Form übergeht. Ein zentraler Baustein dieses Apparats, das Protein SF3B1, ist dafür bekannt, im Bearbeitungszyklus chemisch markiert und entmarkiert zu werden, doch bisher war unklar, welche Rolle diese Markierungen genau spielen und wann sie am wichtigsten sind.
Entdeckung eines angehaltenen Bearbeitungsschritts
Um diese Frage zu untersuchen, verwendeten die Forscher eine kleine Verbindung, die ein Enzym namens CDK11 hemmt, das SF3B1 chemisch markiert. In menschlichen Zellen, die mit diesem Wirkstoff behandelt wurden, isolierten sie an DNA gebundene Spleißosom‑Partikel und bestimmten ihre Proteinbestandteile. Sie entdeckten einen zuvor unbekannten angehaltenen Zustand im Bearbeitungszyklus: ein Komplex, in dem einige Helfergruppen bereits vorhanden sind, während andere noch fehlen. Sie bezeichnen diese arretiert Form als BOTS964. In diesem Zustand hat sich eine frühe Helfergruppe angeschlossen, eine später benötigte Gruppe für die volle Aktivierung ist jedoch noch nicht eingetroffen, was einen spezifischen Kontrollpunkt offenbart, an dem die CDK11‑Aktivität erforderlich ist. 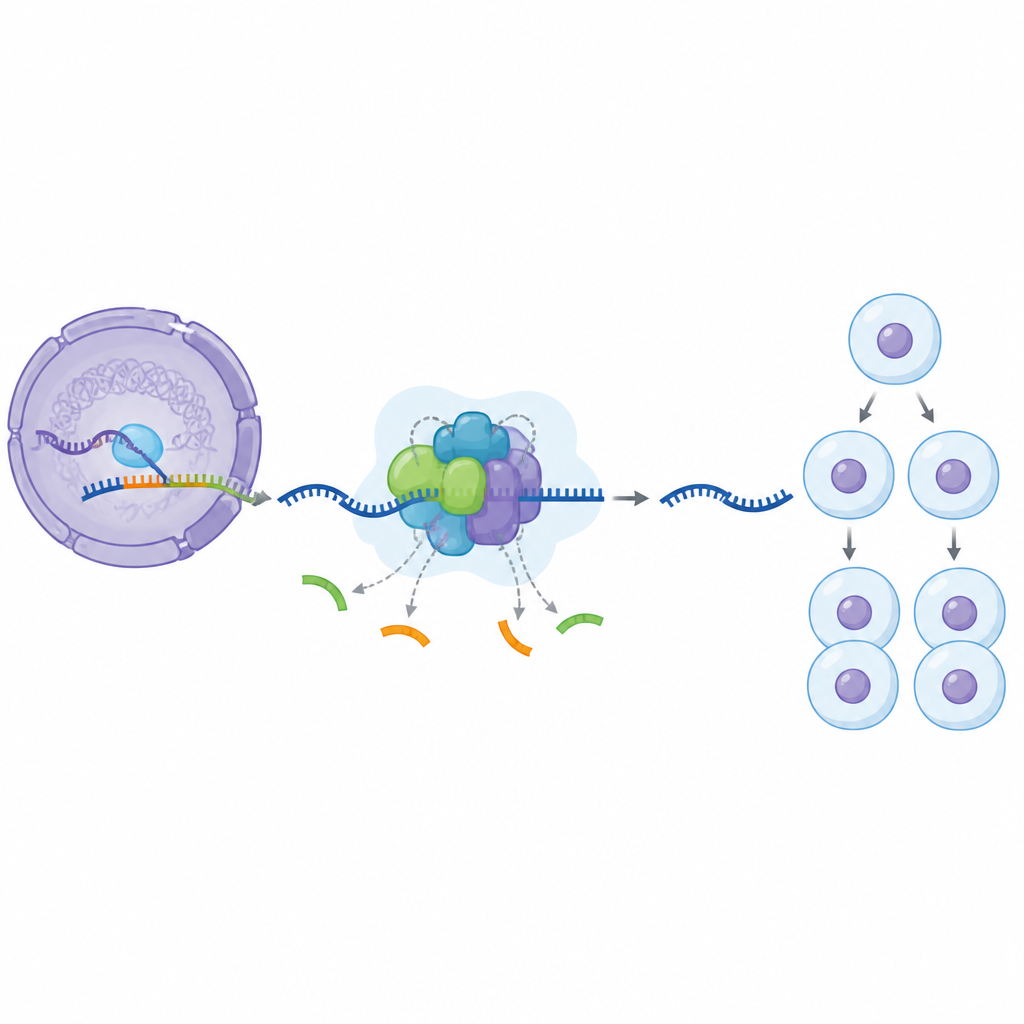
Wie eine Markierung auf SF3B1 das aktive Zentrum formt
Das Team fragte dann, wo die phosphorylierte Form von SF3B1 tatsächlich am RNA sitzt. Mit einer Methode, die Proteine direkt an die RNA‑Basen verknüpft, die sie berühren, kartierten sie die Kontaktpunkte phosphorylierter SF3B1 innerhalb der kleinen RNA‑Stücke, die das Herz des Spleißosoms bilden. Sie fanden, dass markiertes SF3B1 in einer bestimmten Schleife innerhalb der U6‑RNA angereichert ist, einem Bereich, der das katalytische Zentrum formt, in dem Schneiden und Verknüpfen stattfinden. Wenn CDK11 gehemmt wurde, schwächten sich diese Kontakte ab, was darauf hindeutet, dass die Phosphatmarkierung von SF3B1 zur Stabilisierung des gefalteten RNA‑Kerns beiträgt, der für präzises Spleißen nötig ist.
Ein Leserprotein rekrutiert einen wichtigen Helferkomplex
Anschließend suchten die Wissenschaftler nach Proteinen, die bevorzugt an die markierte Form von SF3B1 binden. Sie identifizierten SNIP1, das Teil einer dreikomponentigen Helfergruppe namens RES‑Komplex ist, die dafür bekannt ist, fehlerhafte RNA‑Botschaften am Entweichen aus dem Zellkern zu hindern. SNIP1 trägt eine forkhead‑associated‑Domäne, eine Tasche, die spezifische Phosphatmarken erkennt. Biochemische Tests und strukturelle Modellierung zeigten, dass diese Tasche mehrere phosphorylierte Stellen im flexiblen Schwanz von SF3B1 anbindet. Diese Wechselwirkung hilft, den gesamten RES‑Komplex am Spleißosom zu rekrutieren und zu verankern, genau in dem Moment, in dem es katalytisch aktiv wird, und sorgt so für einen reibungslosen Ablauf des Aktivierungsschritts.
Wenn der Leser fehlt oder geschädigt ist
Um zu sehen, was passiert, wenn SNIP1 plötzlich entfernt wird, erzeugte das Team Zellen, in denen SNIP1 rasch abgebaut werden kann. Innerhalb weniger Stunden zeigten viele Gene weitverbreitete Retention von Introns, was auf einen allgemeinen Zusammenbruch des RNA‑Spleißens hindeutete. Das Muster der Defekte stimmte eng mit demjenigen überein, das bei Hemmung von CDK11 beobachtet wird, was hervorhebt, dass beide Proteine gemeinsam in derselben Phase der Bearbeitung wirken. Ohne SNIP1 kann sich der Großteil des RES‑Komplexes nicht am Spleißosom anschließen, und SF3B1 wird von CDK11 übermäßig markiert, was die Idee stützt, dass die richtige Rekrutierung von SNIP1 von korrekt phosphorylierter SF3B1 abhängt.
Verbindungen zu einer menschlichen Gehirnerkrankung
Schließlich untersuchten die Forscher subtile Veränderungen in der SNIP1‑Tasche, darunter eine E366G‑Mutation, die zuvor bei Menschen aus einer Gemeinschaft mit einer neuro‑entwicklungsbedingten Störung gefunden wurde. Mutante SNIP1‑Proteine bindeten weniger gut an markiertes SF3B1, waren weniger stark mit dem aktiven Spleißosom assoziiert und konnten das Spleißen und Zellwachstum weniger gut retten, wenn das native SNIP1 entfernt wurde. Andere künstliche Mutationen, die diese Interaktion weiter schwächten, verursachten noch stärkere Defekte. Zusammen stützen diese Ergebnisse ein Modell, in dem CDK11 zuerst SF3B1 markiert, markiertes SF3B1 dann SNIP1 und den RES‑Komplex rekrutiert, und diese Ereigniskette das RNA‑katalytische Zentrum stabilisiert und das Spleißen effizient hält. Das Stören eines Glieds dieser Kette, einschließlich krankheitsassoziierter Veränderungen in SNIP1, kann die RNA‑Verarbeitung und die normale Zellfunktion beeinträchtigen. 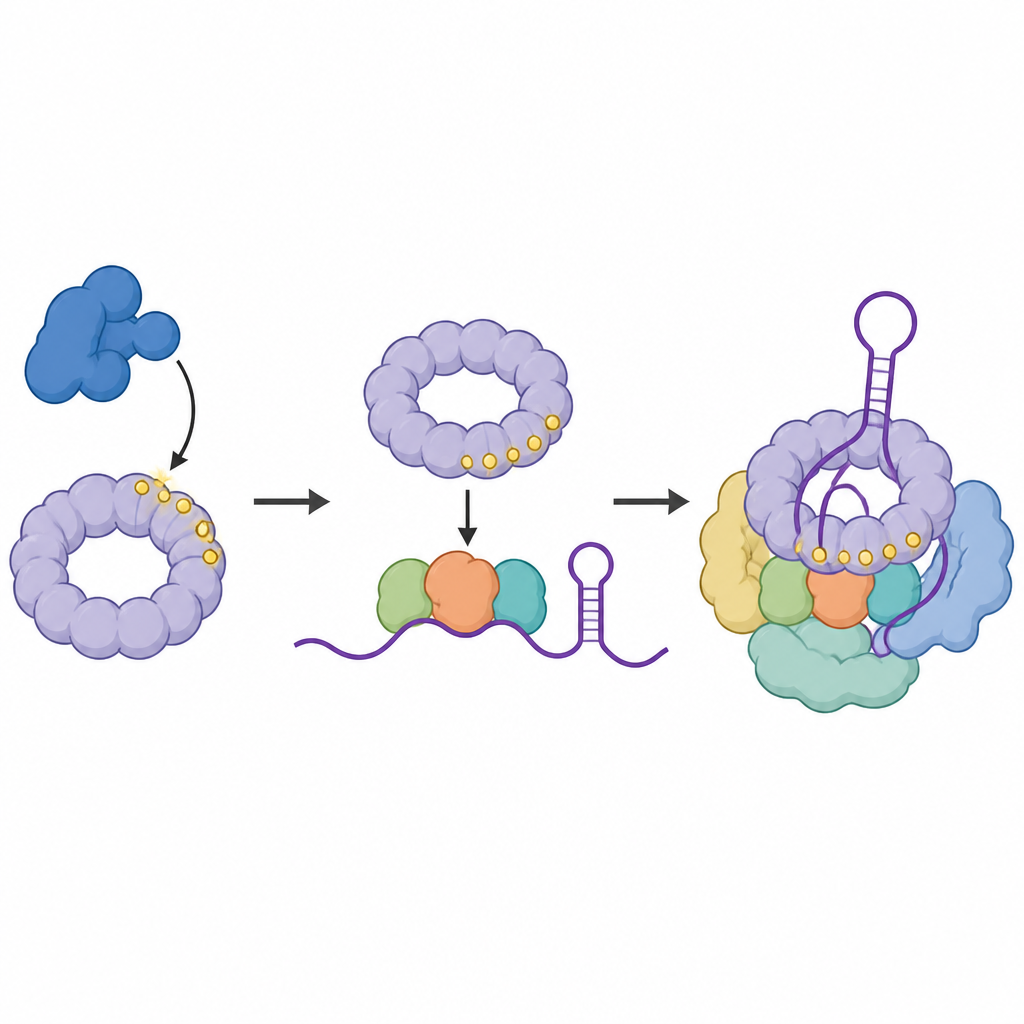
Zitation: Gajdušková, P., Ruiz de Los Mozos, I., Hluchý, M. et al. Phosphorylation of SF3B1 by CDK11 orchestrates spliceosome activation via SNIP1-dependent RES complex recruitment. Nat Commun 17, 4577 (2026). https://doi.org/10.1038/s41467-026-71119-2
Schlüsselwörter: RNA‑Spleißen, Spleißosom, SF3B1, CDK11, SNIP1