Clear Sky Science · nl
Fosforylering van SF3B1 door CDK11 orkestreert activatie van het spliceosoom via SNIP1-afhankelijke rekrutering van het RES-complex
Hoe cellen hun genetische boodschappen bewerken
Elke cel moet ruwe genetische boodschappen zorgvuldig bewerken voordat ze in eiwitten worden omgezet. Wanneer deze bewerking misgaat, kan dat bijdragen aan kanker en hersenaandoeningen. Deze studie onthult hoe een kleine groep eiwitten in onze cellen samenwerkt als een timing-schakelaar om te waarborgen dat RNA-boodschappen correct worden verwerkt, en hoe een enkele erfelijke verandering in een van deze eiwitten deze verfijnde controle kan verstoren.
De cellulaire bewerkingsmachine
Onze genen worden eerst gekopieerd naar lange RNA-strengen die nuttige stukken bevatten, vermengd met extra segmenten die verwijderd moeten worden. Een enorm cellulair complex, het spliceosoom, voert dit knippen en plakken uit. Het bouwt zich in fasen op, waarbij vele eiwitcomponenten worden toegevoegd en losgelaten naarmate het van een vroege rusttoestand naar een volledig actieve vorm gaat. Een kernonderdeel van deze machine, een eiwit genaamd SF3B1, is bekend als chemisch voorzien en ontdaan van tags tijdens de bewerkingscyclus, maar tot nu toe begrepen wetenschappers niet volledig wat deze tags doen of wanneer ze precies het belangrijkst zijn.
Een gepauzeerde bewerkingsstap ontdekken
Om deze vraag te onderzoeken, gebruikten de onderzoekers een klein molecuul dat een enzym genaamd CDK11 blokkeert, dat fosfaatgroepen aan SF3B1 toevoegt. In menselijke cellen die met deze stof werden behandeld, isoleerden ze spliceosoomdeeltjes die aan DNA vastzaten en bepaalden hun eiwitsamenstelling. Ze ontdekten een eerder onbekende gepauzeerde toestand in de bewerkingscyclus: een complex dat sommige hulp-eiwitgroepen al op hun plaats heeft terwijl andere nog ontbreken. Ze noemen deze vastgelopen vorm BOTS964. In deze toestand is een vroege hulpgroep aangesloten, maar een latere groep die normaal voor volledige activatie nodig is, is nog niet gearriveerd, wat wijst op een specifiek controlepunt waarbij CDK11-activiteit vereist is. 
Hoe een tag op SF3B1 de actieve plek vormt
Het team vroeg zich vervolgens af waar de getagde versie van SF3B1 precies op RNA zit. Met een techniek die eiwitten direct aan de RNA-basen koppelt die ze aanraken, brachten ze de contactpunten van gefosforyleerd SF3B1 in kaart binnen de kleine RNA-stukjes die het hart van het spliceosoom vormen. Ze vonden dat getagd SF3B1 verrijkt is op een specifieke lus binnen U6-RNA, een regio die helpt het katalytische centrum te vormen waar knippen en plakken plaatsvinden. Wanneer CDK11 werd geblokkeerd, verzwakten deze contacten, wat suggereert dat het toevoegen van fosfaatgroepen aan SF3B1 bijdraagt aan het stabiliseren van het gevouwen RNA-centrum dat nodig is voor nauwkeurige splitsing.
Een lezer-eiwit rekruteert een cruciaal hulpcomplex
Vervolgens zochten de wetenschappers naar eiwitten die de voorkeur geven aan binding aan de getagde vorm van SF3B1. Ze identificeerden SNIP1, onderdeel van een drie-eiwits hulpcomplex genaamd het RES-complex, dat bekendstaat om te voorkomen dat defecte RNA-boodschappen de kern verlaten. SNIP1 draagt een forkhead-associated (FHA) domein, een pocket die specifieke fosfaatmerken herkent. Biochemische testen en structurele modellering toonden aan dat deze pocket meerdere gefosforyleerde plekken binnen de flexibele staart van SF3B1 aangrijpt. Deze interactie helpt het volledige RES-complex te rekruteren en te verankeren op het spliceosoom juist op het moment dat het katalytisch actief wordt, waardoor een soepele voortgang door de activatiestap wordt gegarandeerd.
Wanneer de lezer ontbreekt of beschadigd is
Om te zien wat er gebeurt als SNIP1 plotseling verdwijnt, maakten de onderzoekers cellen waarin SNIP1 snel kan worden afgebroken. Binnen enkele uren na degradatie vertoonden veel genen wijdverspreide retentie van introns, wat wijst op een brede ineenstorting van RNA-splitsing. Het patroon van defecten kwam nauw overeen met wat wordt gezien wanneer CDK11 wordt geremd, wat benadrukt dat beide eiwitten samenwerken in dezelfde fase van de bewerking. Zonder SNIP1 slaagt het grootste deel van het RES-complex er niet in het spliceosoom te betreden, en wordt SF3B1 overmatig gefosforyleerd door CDK11, wat het idee versterkt dat correcte rekrutering van SNIP1 afhangt van een juist gefosforyleerd SF3B1.
Verbonden met een menselijke hersenaandoening
Tenslotte onderzochten de onderzoekers subtiele veranderingen in de SNIP1-pocket, waaronder een E366G-mutatie die eerder is gevonden bij mensen uit een gemeenschap met een neuro-ontwikkelingsstoornis. Mutante SNIP1-eiwitten binden minder goed aan getagd SF3B1, associëren minder sterk met het actieve spliceosoom en waren minder in staat om splitsing en celgroei te herstellen wanneer het endogene SNIP1 werd verwijderd. Andere kunstmatige mutaties die deze interactie verder verzwakten veroorzaakten nog sterkere defecten. Gezamenlijk ondersteunen deze resultaten een model waarin CDK11 eerst SF3B1 tagt, getagd SF3B1 vervolgens SNIP1 en het RES-complex rekruteert, en deze keten van gebeurtenissen het RNA-katalytische centrum stabiliseert en de efficiëntie van splitsing bewaakt. Het verstoren van een schakel in deze keten, inclusief ziekte-geassocieerde veranderingen in SNIP1, kan RNA-processing en normale celfunctie aantasten. 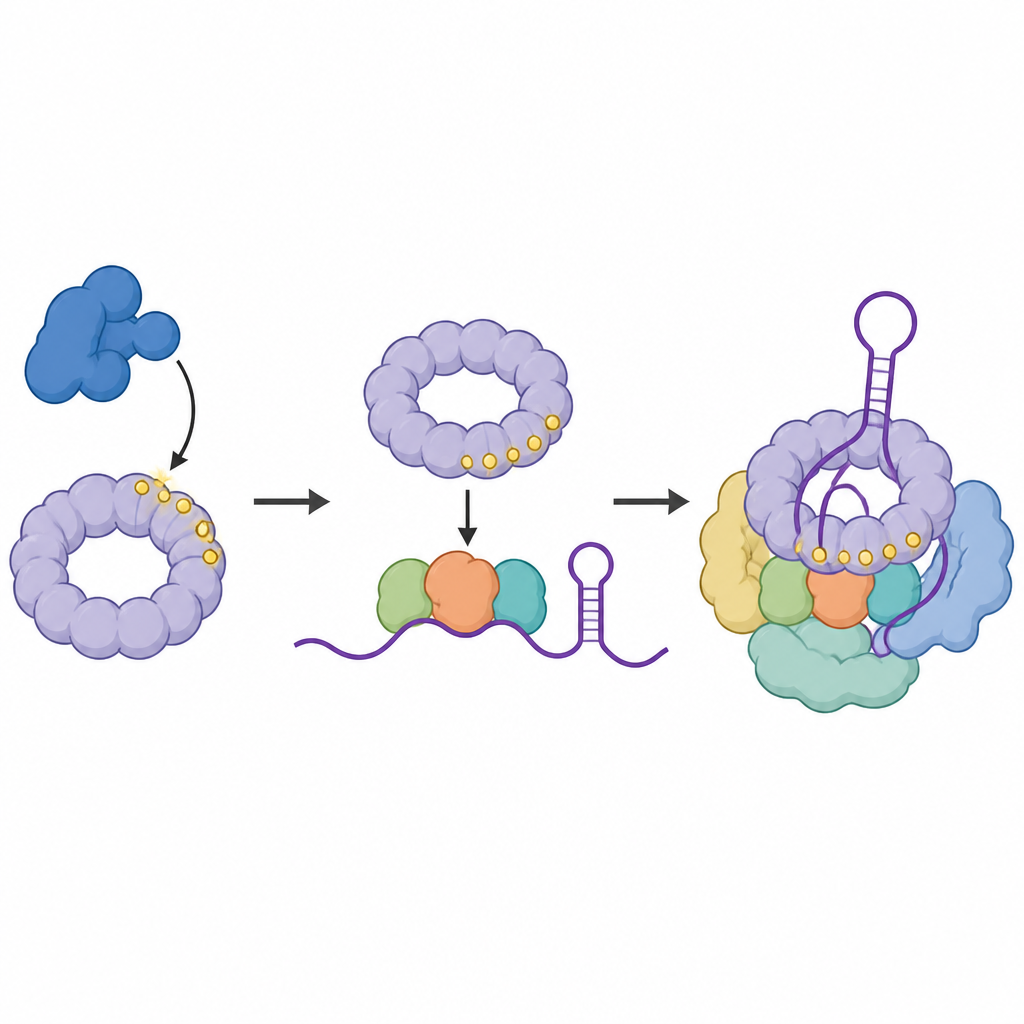
Bronvermelding: Gajdušková, P., Ruiz de Los Mozos, I., Hluchý, M. et al. Phosphorylation of SF3B1 by CDK11 orchestrates spliceosome activation via SNIP1-dependent RES complex recruitment. Nat Commun 17, 4577 (2026). https://doi.org/10.1038/s41467-026-71119-2
Trefwoorden: RNA-splitsing, spliceosoom, SF3B1, CDK11, SNIP1