Clear Sky Science · pl
Fosforylacja SF3B1 przez CDK11 orkiestruje aktywację spliceosomu poprzez rekrutację kompleksu RES zależną od SNIP1
Jak komórki edytują swoje genetyczne komunikaty
Każda komórka musi starannie edytować surowe komunikaty genetyczne, zanim przekształci je w białka. Gdy ta edycja zawiedzie, może przyczyniać się do powstawania nowotworów i zaburzeń mózgu. W tym badaniu ujawniono, jak niewielka grupa białek w komórkach współdziała jako przełącznik czasowy, by zapewnić poprawne przetwarzanie komunikatów RNA, oraz jak pojedyncza dziedziczna zmiana w jednym z tych białek może zaburzyć tę delikatną kontrolę.
Komórkowa maszyna do edycji
Nasze geny są najpierw przepisywane na długie nici RNA, które zawierają użyteczne fragmenty wymieszane z dodatkowymi odcinkami, które należy usunąć. Ogromna komórkowa maszyna zwana spliceosomem wykonuje to cięcie i łączenie. Składa się ona etapami, dodając i uwalniając wiele składników białkowych, gdy przechodzi ze wczesnego stanu spoczynkowego do w pełni aktywnej formy. Jednym z kluczowych elementów tej maszyny jest białko SF3B1, o którym wiadomo, że w cyklu edycyjnym ulega chemicznemu oznaczaniu i odznaczaniu, lecz do tej pory naukowcy nie rozumieli w pełni, co te znaczniki robią i kiedy są najistotniejsze.
Odnalezienie zatrzymanego etapu edycji
Aby zbadać to zagadnienie, badacze zastosowali małą cząsteczkę blokującą enzym o nazwie CDK11, który przyłącza chemiczne znaczniki do SF3B1. W komórkach ludzkich traktowanych tym związkiem wyizolowali cząstki spliceosomu związane z DNA i zmierzyli ich skład białkowy. Odkryli wcześniej nieznany zatrzymany stan w cyklu edycyjnym: kompleks, który ma już zmontowane niektóre grupy pomocnicze, podczas gdy inne wciąż są nieobecne. Nazwali tę zahamowaną formę BOTS964. W tym stanie dołączyła wczesna grupa pomocnicza, lecz późniejsza, zwykle niezbędna do pełnej aktywacji, jeszcze nie nadeszła, co ujawnia konkretny punkt kontrolny, w którym wymagana jest aktywność CDK11. 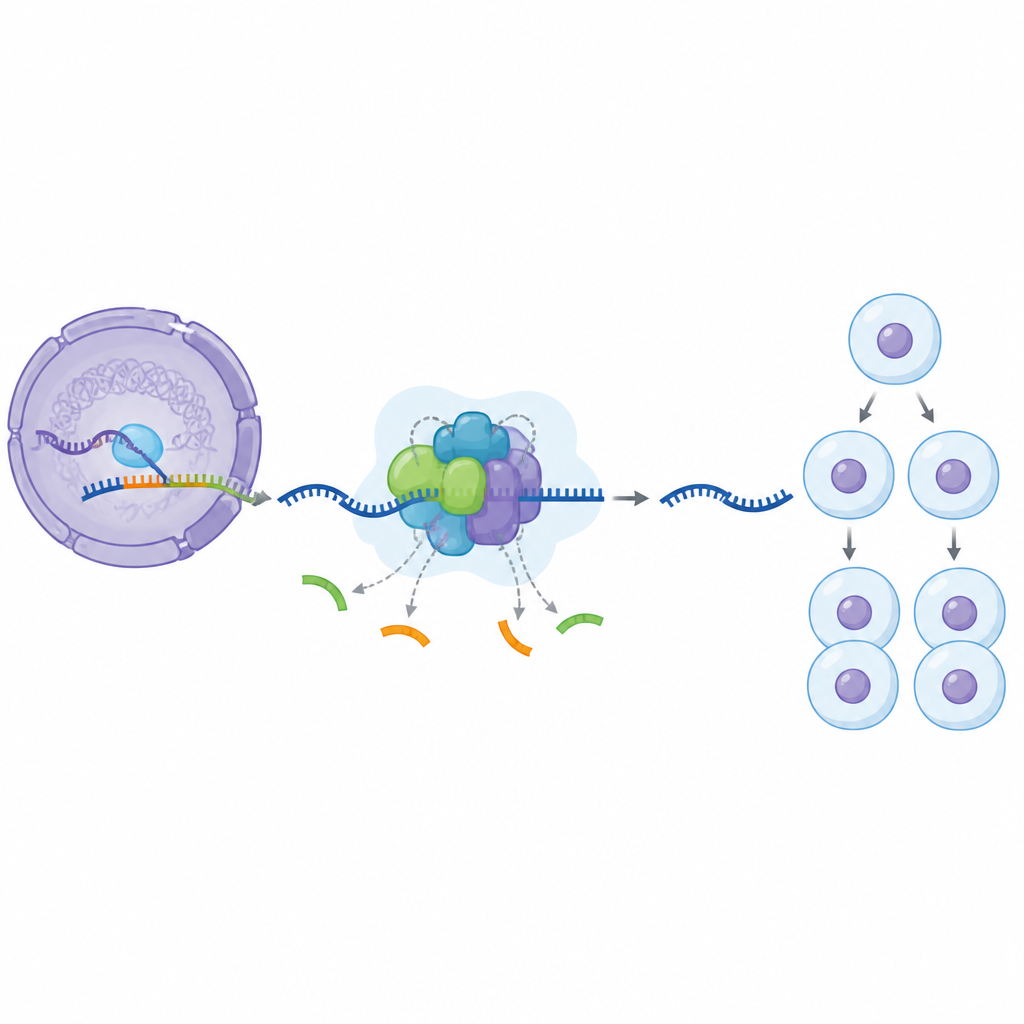
Jak znacznik na SF3B1 kształtuje miejsce aktywne
Zespół zapytał następnie, gdzie wersja SF3B1 z przyłączonymi znacznikami faktycznie znajduje się na RNA. Stosując technikę krzyżowania białek bezpośrednio z zasadami RNA, które dotykają, zmapowali punkty kontaktu sfosforylowanego SF3B1 w obrębie małych fragmentów RNA tworzących rdzeń spliceosomu. Odkryli, że znakowany SF3B1 jest wzbogacony w określonej pętli w RNA U6 — regionie pomagającym ukształtować katalityczne centrum, gdzie zachodzą cięcie i łączenie. Gdy CDK11 był zablokowany, te kontakty osłabły, co sugeruje, że przyłączanie reszt fosforanowych do SF3B1 pomaga stabilizować złożone centrum RNA potrzebne do precyzyjnego splicingu.
Białko-odczytywacz rekrutuje kluczowy kompleks pomocniczy
Następnie naukowcy poszukali białek, które wolą wiązać znakowaną formę SF3B1. Zidentyfikowali SNIP1, część trzybiałkowego zespołu pomocniczego zwanego kompleksem RES, znanego z zapobiegania wydostawaniu się błędnych komunikatów RNA z jądra komórkowego. SNIP1 posiada domenę forkhead-associated, kieszeń rozpoznającą specyficzne znaki fosforanowe. Badania biochemiczne i modelowanie strukturalne wykazały, że ta kieszeń angażuje wiele sfosforylowanych miejsc w elastycznym końcu SF3B1. Ta interakcja pomaga rekrutować i zakotwiczać cały kompleks RES na spliceosomie w momencie jego przejścia w formę katalitycznie aktywną, zapewniając płynne przejście przez etap aktywacji.
Gdy odczytywacz jest nieobecny lub uszkodzony
Aby zobaczyć, co się dzieje po nagłym usunięciu SNIP1, zespół skonstruował komórki, w których SNIP1 można szybko degradować. W ciągu godzin od jego wyczerpania wiele genów wykazywało szerokie zatrzymywanie intronów, wskazując na masywny zanik poprawnego splicingu RNA. Wzór defektów ściśle odpowiadał temu, co obserwuje się przy hamowaniu CDK11, podkreślając, że oba białka działają wspólnie na tym samym etapie edycji. Bez SNIP1 większość kompleksu RES nie dołącza do spliceosomu, a SF3B1 staje się nadmiernie znakowany przez CDK11, co wzmacnia przekonanie, że prawidłowa rekrutacja SNIP1 zależy od właściwej fosforylacji SF3B1.
Powiązania z ludzkim zaburzeniem mózgowym
Na koniec badacze przeanalizowali subtelne zmiany w kieszeni SNIP1, w tym mutację E366G wykrytą wcześniej u osób z jednej społeczności z zaburzeniem neurorozwojowym. Zmutowane białka SNIP1 wiązały się słabiej ze znakowanym SF3B1, słabiej łączyły się z aktywnym spliceosomem i były mniej zdolne do przywrócenia splicingu i wzrostu komórek po usunięciu natywnego SNIP1. Inne sztuczne mutacje, które dodatkowo osłabiały tę interakcję, powodowały jeszcze silniejsze defekty. Razem te wyniki wspierają model, w którym CDK11 najpierw znakuję SF3B1, znakowany SF3B1 rekrutuje następnie SNIP1 i kompleks RES, a ten ciąg zdarzeń stabilizuje katalityczne centrum RNA i utrzymuje wydajny splicing. Zakłócenie któregokolwiek ogniwa tej sekwencji, w tym zmian związanych z chorobą w SNIP1, może upośledzić przetwarzanie RNA i prawidłową funkcję komórkową. 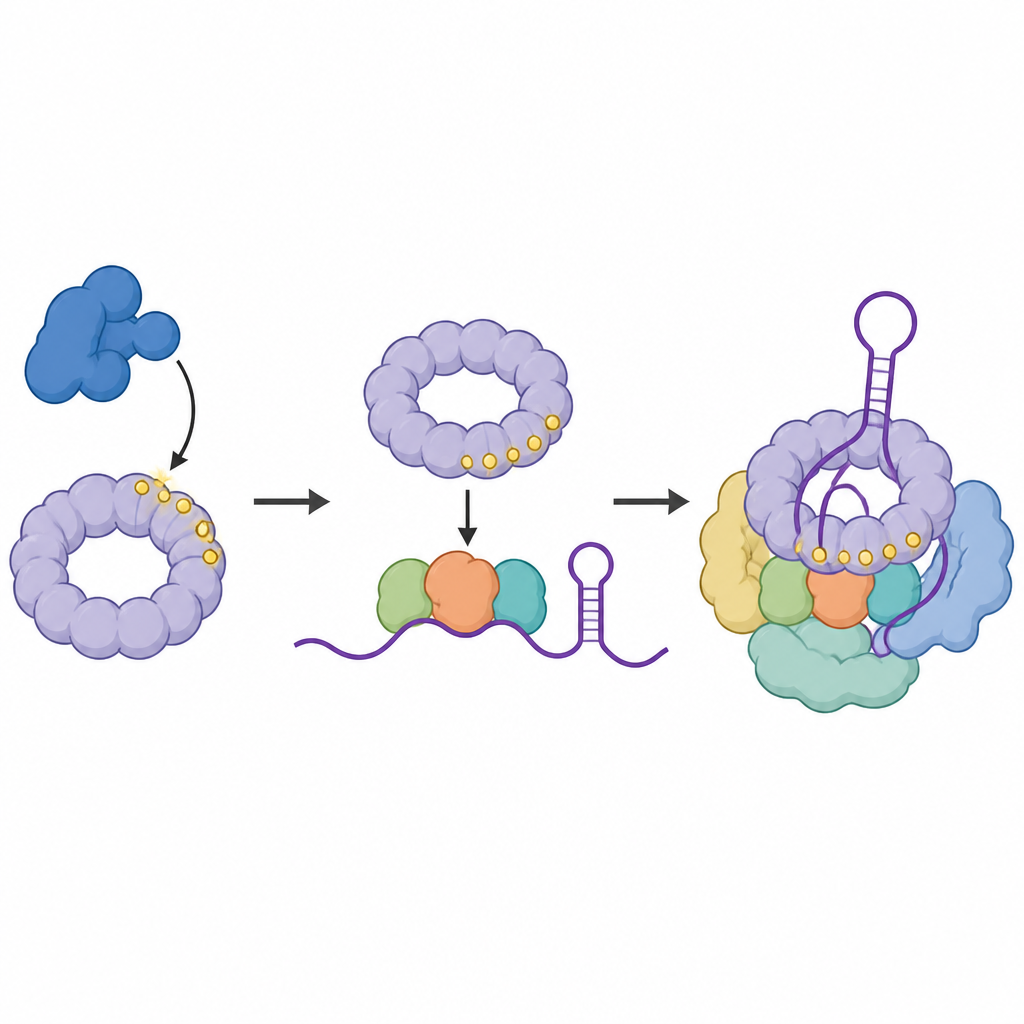
Cytowanie: Gajdušková, P., Ruiz de Los Mozos, I., Hluchý, M. et al. Phosphorylation of SF3B1 by CDK11 orchestrates spliceosome activation via SNIP1-dependent RES complex recruitment. Nat Commun 17, 4577 (2026). https://doi.org/10.1038/s41467-026-71119-2
Słowa kluczowe: splicing RNA, spliceosom, SF3B1, CDK11, SNIP1