Clear Sky Science · pt
Uma anidrase carbônica em células-guarda se liga e regula SLAC1 separadamente de sua atividade catalítica
Por que poros minúsculos nas folhas importam
Cada folha é pontilhada por poros microscópicos que funcionam como válvulas ajustáveis, trocando água por dióxido de carbono. Essas válvulas, chamadas estômatos, precisam se abrir para permitir a entrada de CO2 para a fotossíntese, mas fechar rapidamente para evitar que a planta resseque. Como os estômatos detectam e reagem em tempo real às mudanças nos níveis de dióxido de carbono tem sido um quebra‑cabeça de longa data. Este estudo revela uma peça chave desse mecanismo, mostrando como duas proteínas nas células que cercam cada poro trabalham em conjunto para ajustar com precisão a perda de água e o crescimento.
Válvulas foliares que equilibram ar e água
Os estômatos consistem em pares de células‑guarda que incham para abrir ou murcham para fechar ao mover sais e água para dentro e fora. Quando o ar ao redor ou os espaços aéreos dentro da folha contêm mais dióxido de carbono, os estômatos tendem a se fechar. Isso protege a planta de desperdiçar água quando há CO2 extra disponível. Uma via importante para liberar íons carregados negativamente durante o fechamento é um canal de membrana chamado SLAC1. Trabalhos anteriores sugeriram que uma família de enzimas conhecidas como anidrases carbônicas, que normalmente aceleram a conversão entre dióxido de carbono e bicarbonato em água, de alguma forma ajudava a ligar o SLAC1. Mas não estava claro se essas enzimas atuavam apenas como conversores químicos ou também como botões de controle diretos do canal.
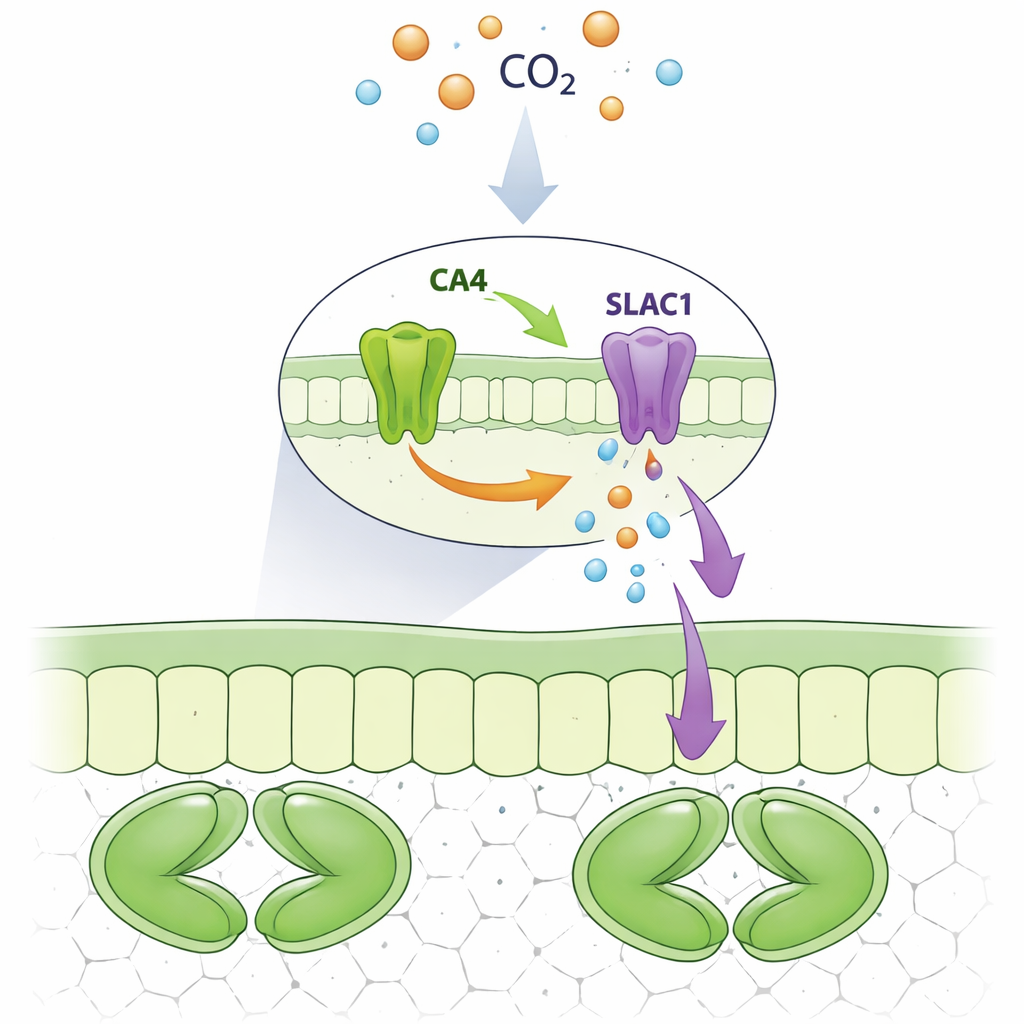
Uma parceria proteica no poro
Os pesquisadores se concentraram em uma anidrase carbônica específica, CA4, que é abundante em células‑guarda. Usando células de levedura e imagem fluorescente em células vegetais, eles mostraram que SLAC1 forma um complexo físico com CA4, mas não com enzimas intimamente relacionadas CA1 ou CA3. Ambas as versões de CA4 encontradas em plantas, uma ligada a membranas e outra mais livre no fluido celular, puderam se ligar ao SLAC1. A equipe então alterou sistematicamente aminoácidos individuais em CA4 para ver quais eram necessários para esse aperto de mão. Eles descobriram um pequeno trecho superficial de CA4, centrado em alguns resíduos específicos distantes do sítio de reação química, que é essencial para a ligação ao SLAC1. Mutações nesse motivo romperam a interação enquanto deixavam intacta a atividade catalítica da enzima.
Separando química de controle
Ao separar o papel de ligação de CA4 do seu papel catalítico, os autores investigaram como cada um afetava o canal SLAC1. Em ovos de rã modificados para produzir as proteínas vegetais, a adição de CA4 normal aumentou a corrente iônica de SLAC1, enquanto formas mutantes de CA4 que não conseguiam mais se ligar ao canal não promoveram esse aumento. De forma notável, um mutante de CA4 que havia perdido a atividade catalítica mas ainda se ligava ao SLAC1 foi capaz de aumentar a corrente. Em células‑guarda de folhas de Arabidopsis, CA4 normal restaurou um aumento robusto na atividade de SLAC1 dependente de dióxido de carbono. Em contraste, plantas que expressavam CA4 com defeito de ligação mostraram pouco ou nenhum aumento na atividade do canal mesmo com CO2 elevado, apesar de alguns desses mutantes ainda realizarem a química normalmente. Isso mostra que o contato físico direto de CA4 com SLAC1, e não apenas sua capacidade de processar CO2, é o que ajusta a resposta do canal.
Do canal único ao desempenho da planta inteira
A equipe então acompanhou as consequências de interromper a ligação CA4–SLAC1 na escala de folhas e plantas inteiras. Em plantas nas quais CA4 não conseguia se prender ao SLAC1, os estômatos fecharam e reabriram muito mais lentamente após o aumento ou a normalização dos níveis de dióxido de carbono. Modelos computacionais previram esse comportamento lento e sugeriram que ele tornaria as plantas menos eficientes no uso da água. Experimentos sob luz flutuante controlada confirmaram essas previsões: plantas com CA4 com ligação prejudicada apresentaram rosetas menores, menor peso seco e eficiência no uso da água significativamente pior do que plantas com CA4 normal ou com CA4 deficiente em catálise mas competente em ligação. Importante, a maquinaria fotossintética em si funcionou igualmente bem, indicando que a penalidade de crescimento decorreu principalmente de movimentos estomatais fora de tempo em vez de uma fixação de carbono defeituosa.
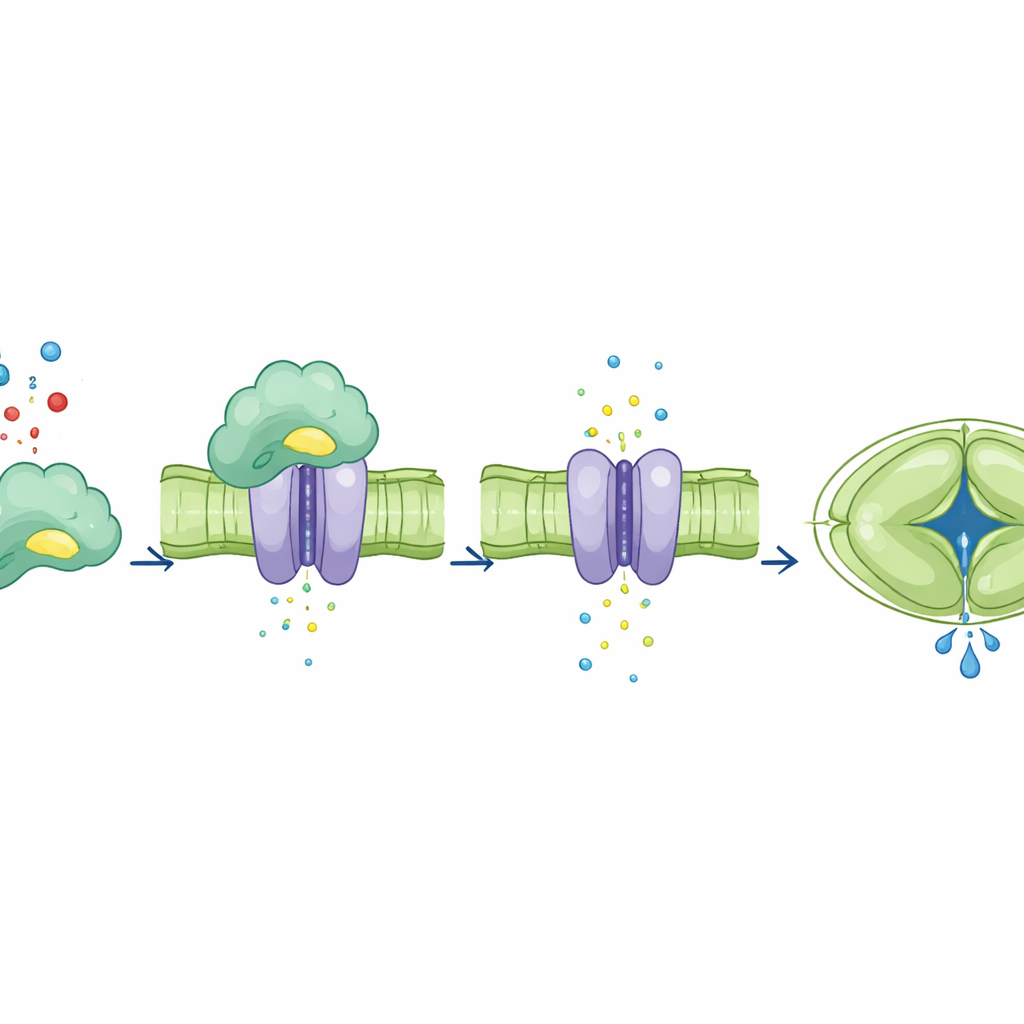
O que isso significa para culturas do futuro
Em conjunto, os resultados revelam CA4 como um parceiro‑sensor que se prende ao canal SLAC1 em células‑guarda e ajusta diretamente sua atividade em resposta ao CO2 próximo ao ambiente. Essa ligação usa um motivo estrutural distinto do centro de reação química da enzima, provando que seu papel regulatório pode ser desvinculado de seu papel catalítico. Ao afiar a rapidez com que os estômatos se abrem e se fecham, essa parceria proteica ajuda as plantas a equilibrar melhor a captação de carbono e a economia de água, especialmente sob condições de luz natural que mudam de minuto a minuto. Na prática, o trabalho aponta para novos alvos moleculares para o melhoramento ou engenharia de culturas cujas válvulas foliares respondam com mais agilidade, potencialmente aumentando rendimentos enquanto usam menos água em um mundo mais quente e seco.
Citação: Xia, L., Alvim, J.C., Nguyen, TH. et al. A guard cell carbonic anhydrase binds and regulates SLAC1 separate from its catalytic activity. Nat Commun 17, 3911 (2026). https://doi.org/10.1038/s41467-026-70596-9
Palavras-chave: estômatos, sensoriamento de dióxido de carbono, células-guarda, eficiência no uso da água, canais iônicos