Clear Sky Science · nl
Een koolzuuranhydrase in sluitcellen bindt SLAC1 en regelt het onafhankelijk van zijn katalytische activiteit
Waarom kleine bladspleetjes ertoe doen
Elk blad is bezaaid met microscopische openingen die werken als instelbare kleppen en water ruilen voor kooldioxide. Deze kleppen, stomata genoemd, moeten openstaan om kooldioxide voor fotosynthese binnen te laten, maar sluiten snel om te voorkomen dat planten uitdrogen. Hoe stomata realtime veranderingen in kooldioxide waarnemen en erop reageren is lange tijd een raadsel geweest. Deze studie onthult een belangrijk deel van dat mechanisme en laat zien hoe twee eiwitten in de cellen rond elke opening samenwerken om waterverlies en groei fijn af te stemmen.
Bladkleppen die lucht en water in evenwicht houden
Stomata bestaan uit paren sluitcellen die door het verplaatsen van zouten en water openzwellen of sluiten. Wanneer de omliggende lucht of de luchtruimten in het blad meer kooldioxide bevatten, hebben stomata de neiging te sluiten. Dit beschermt de plant tegen onnodig waterverlies wanneer er al extra kooldioxide beschikbaar is. Een belangrijke route voor het vrijlaten van negatief geladen ionen tijdens het sluiten is een membraankanaal genaamd SLAC1. Eerder werk gaf al aan dat een familie enzymen, koolzuuranhydrases geheten, die normaal gesproken de omzetting tussen kooldioxide en bicarbonaat in water versnellen, op de één of andere manier hielpen SLAC1 te activeren. Maar het was onduidelijk of deze enzymen alleen als chemische omzetters werkten of dat ze ook als directe bedieningsknoppen voor het kanaal fungeerden.
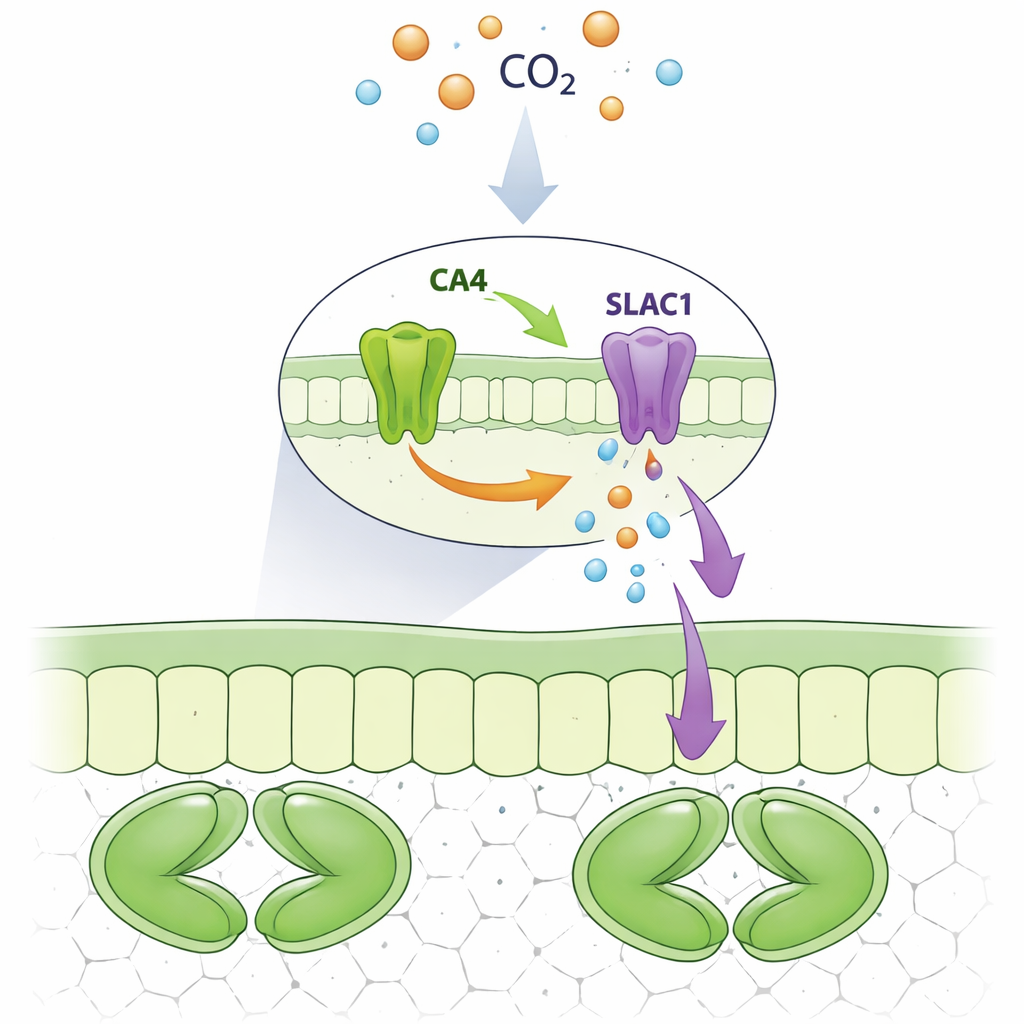
Een eiwitpartnerschap bij de opening
De onderzoekers concentreerden zich op een specifieke koolzuuranhydrase, CA4, die overvloedig aanwezig is in sluitcellen. Met behulp van gistcellen en fluorescentiebeeldvorming in plantencellen lieten ze zien dat SLAC1 een fysiek complex vormt met CA4, maar niet met nauw verwante enzymen CA1 of CA3. Beide vormen van CA4 die in planten voorkomen — één aan membranen gebonden en één vrij in het celvocht — konden aan SLAC1 binden. Het team wijzigde daarna systematisch individuele aminozuren in CA4 om te achterhalen welke nodig waren voor deze handdruk. Ze ontdekten een kort oppervlaktedeeletje van CA4, gecentreerd rond een paar specifieke residuen ver van de chemische actieve plaats, dat essentieel is voor het binden van SLAC1. Mutaties in dit motief verbraken de interactie terwijl de katalytische activiteit van het enzym intact bleef.
Chemie scheiden van controle
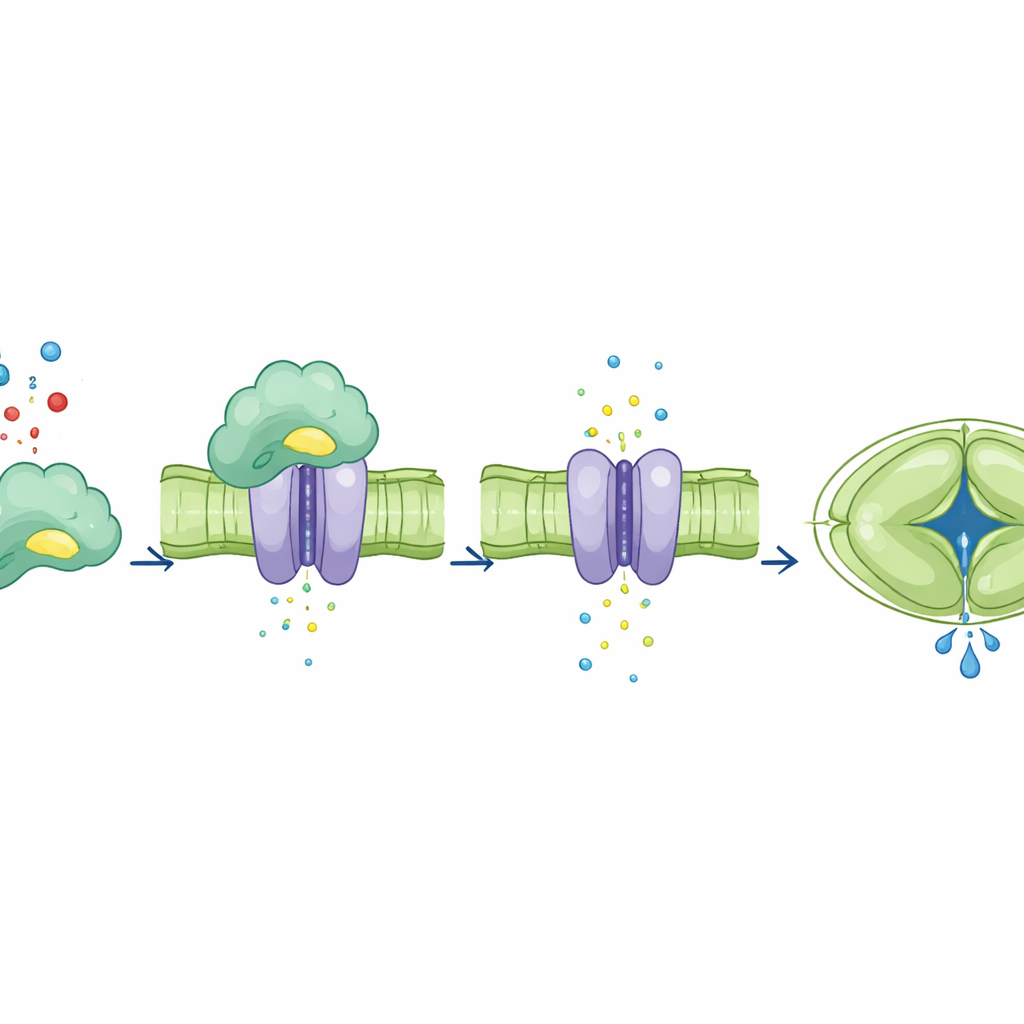
Nadat ze CA4’s bindende rol hadden gescheiden van zijn katalytische rol, onderzochten de auteurs hoe elk de SLAC1‑kanaal beïnvloedde. In kikker‑eieren die waren getransformeerd om de planteiwitten te produceren, verhoogde toevoeging van normale CA4 de ionenstroom van SLAC1, terwijl gemuteerde vormen van CA4 die het kanaal niet meer konden binden dat niet deden. Opmerkelijk was dat een CA4‑mutant die zijn katalytische activiteit had verloren maar nog steeds aan SLAC1 bond, de stroom nog steeds versterkte. In sluitcellen van Arabidopsis‑bladeren herstelde normale CA4 een sterke, kooldioxide‑afhankelijke toename in SLAC1‑activiteit. Daarentegen vertoonden planten die bindings‑defecte CA4 tot expressie brachten weinig of geen toename in kanaalactiviteit, zelfs bij verhoogd kooldioxide, ondanks dat sommige van deze mutanten nog steeds normale chemie lieten zien. Dit toont aan dat CA4’s directe fysieke contact met SLAC1, en niet alleen zijn vermogen om kooldioxide te verwerken, bepaalt hoe het kanaal reageert.
Van enkele kanalen naar prestaties van de hele plant
Het team volgde vervolgens de gevolgen van het verstoren van CA4–SLAC1‑binding op schaal van hele bladeren en planten. In planten waarin CA4 zich niet aan SLAC1 kon vasthechten, sloten en heropenden stomata veel langzamer nadat de kooldioxideconcentratie was verhoogd of weer normaal werd. Computermodellen voorspelden dit trage gedrag en suggereerden dat het planten minder efficiënt zou maken in hun watergebruik. Experimenten onder gecontroleerd, fluctuerend licht bevestigden deze voorspellingen: planten met binding‑verstoorde CA4 hadden kleinere rozetten, lagere droge massa en significant slechtere watergebruiksefficiëntie dan planten met normale of katalyse‑deficiënte maar binding‑competente CA4. Belangrijk is dat hun fotosynthetische machinerie zelf net zo goed functioneerde, wat aangeeft dat de groeipenalty vooral voortkwam uit verkeerd getimede stomatabewegingen en niet uit defecte koolstoffixatiechemie.
Wat dit betekent voor toekomstige gewassen
Gezamenlijk onthullen de resultaten CA4 als een sensor‑partner die zich vastklampt aan het SLAC1‑kanaal in sluitcellen en de activiteit ervan direct afstemt als reactie op nabij‑omgevingskooldioxide. Deze binding gebruikt een structureel motief dat verschilt van het chemische reactieterrein van het enzym, wat bewijst dat zijn regulatorische rol losgekoppeld kan worden van zijn katalytische rol. Door te verscherpen hoe snel stomata openen en sluiten, helpt dit eiwitpartnerschap planten een betere balans te vinden tussen koolstofopname en waterbesparing, vooral onder natuurlijke lichtomstandigheden die van minuut tot minuut veranderen. In praktische termen wijst dit werk op nieuwe moleculaire doelen voor veredeling of engineering van gewassen waarvan de bladkleppen sneller reageren, wat mogelijk opbrengst kan verhogen terwijl minder water wordt gebruikt in een opwarmende, uitdrogende wereld.
Bronvermelding: Xia, L., Alvim, J.C., Nguyen, TH. et al. A guard cell carbonic anhydrase binds and regulates SLAC1 separate from its catalytic activity. Nat Commun 17, 3911 (2026). https://doi.org/10.1038/s41467-026-70596-9
Trefwoorden: stomata, kooldioxidewaarneming, sluitcellen, watergebruiksefficiëntie, ionkanalen