Clear Sky Science · es
Una anhidrasa carbónica en células guardianas se une y regula SLAC1 separada de su actividad catalítica
Por qué importan los diminutos poros de las hojas
Cada hoja está salpicada de poros microscópicos que actúan como válvulas ajustables, intercambiando agua por dióxido de carbono. Estas válvulas, llamadas estomas, deben abrirse para dejar entrar dióxido de carbono para la fotosíntesis, pero cerrarse rápidamente para evitar que la planta se deshidrate. Cómo los estomas detectan y reaccionan en tiempo real a los cambios en los niveles de dióxido de carbono ha sido un enigma persistente. Este estudio descubre una pieza clave de ese mecanismo, mostrando cómo dos proteínas en las células que rodean cada poro trabajan juntas para afinar la pérdida de agua y el crecimiento.
Válvulas foliares que malaban aire y agua
Los estomas consisten en pares de células guardianas que se hinchan para abrirse o se encogen para cerrarse al mover sales y agua dentro y fuera. Cuando el aire circundante o los espacios de aire dentro de la hoja contienen más dióxido de carbono, los estomas tienden a cerrarse. Esto protege a la planta de desperdiciar agua cuando ya hay dióxido de carbono adicional disponible. Una vía importante para liberar iones cargados negativamente durante el cierre es un canal en la membrana llamado SLAC1. Trabajos anteriores sugirieron que una familia de enzimas conocidas como anhidrasa carbónica, que normalmente aceleran la conversión entre dióxido de carbono y bicarbonato en agua, de algún modo ayudaban a activar SLAC1. Pero no estaba claro si estas enzimas actuaban solo como convertidores químicos o si también funcionaban como botones de control directos para el canal.
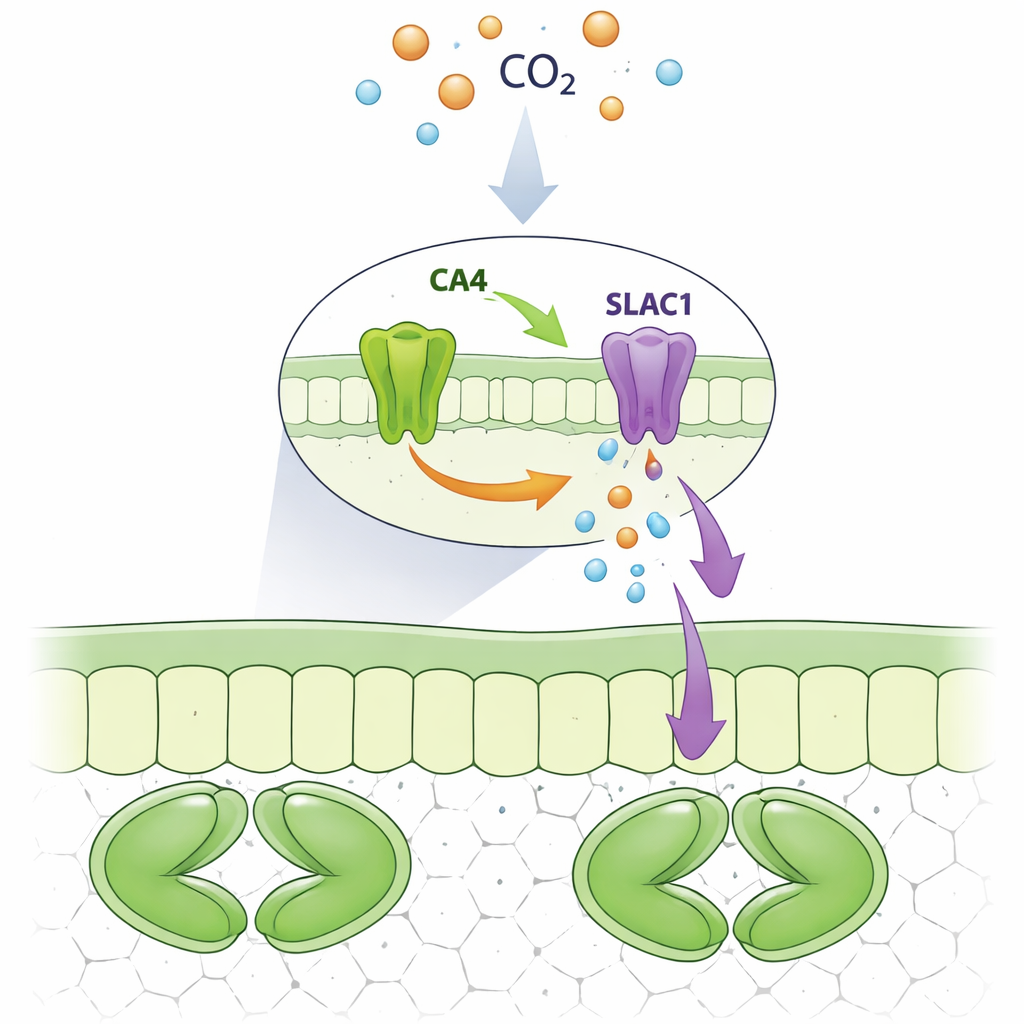
Una asociación proteica en el poro
Los investigadores se centraron en una anhidrasa carbónica particular, CA4, que es abundante en las células guardianas. Usando células de levadura e imágenes fluorescentes en células vegetales, mostraron que SLAC1 forma un complejo físico con CA4, pero no con enzimas estrechamente relacionadas como CA1 o CA3. Ambas versiones de CA4 presentes en las plantas, una unida a membranas y otra más libre en el fluido celular, pudieron unirse a SLAC1. El equipo alteró sistemáticamente aminoácidos individuales en CA4 para ver cuáles eran necesarios para este apretón de manos. Descubrieron un pequeño parche superficial de CA4, centrado en unos pocos residuos específicos alejados del sitio de reacción química, que es esencial para la unión a SLAC1. Mutaciones en este motivo rompieron la interacción mientras dejaban intacta la actividad catalítica de la enzima.
Separando la química del control
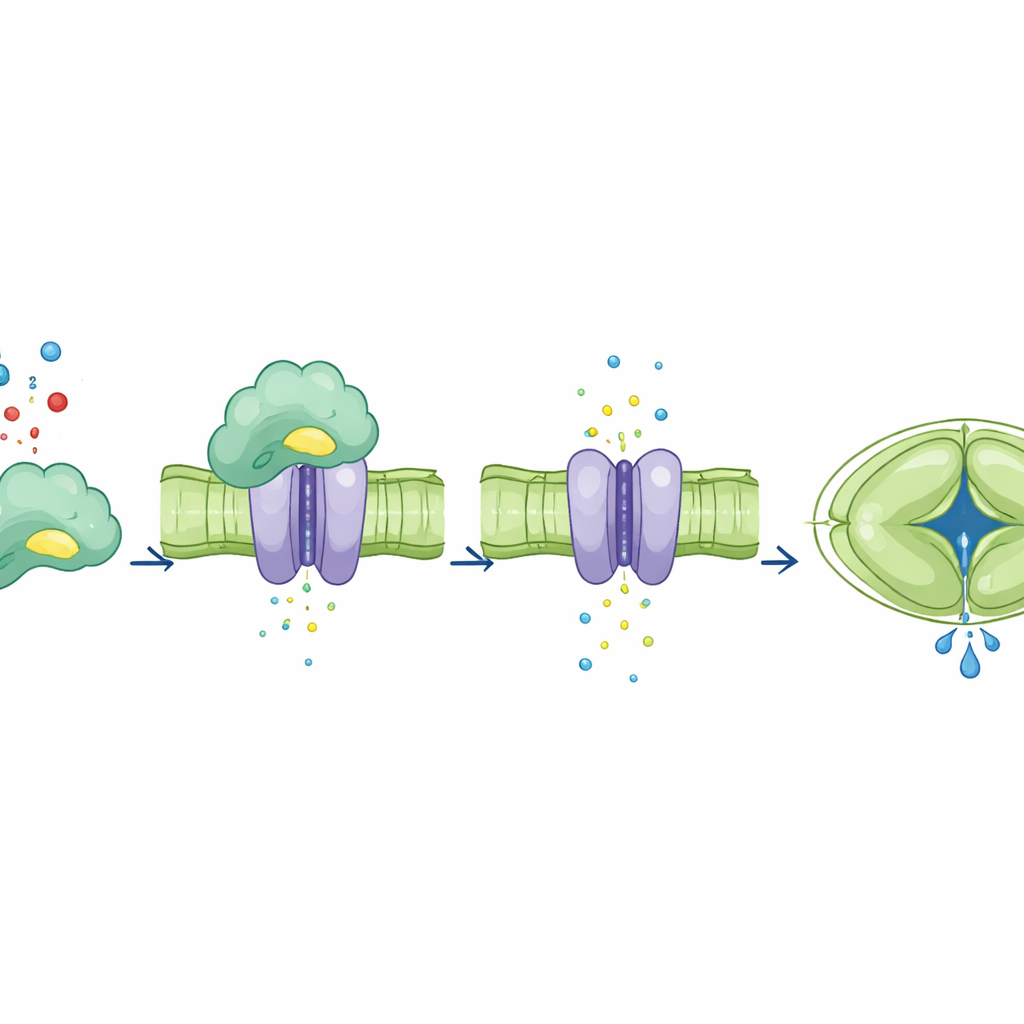
Al haber separado el papel de unión de CA4 de su papel catalítico, los autores preguntaron cómo afectaba cada uno al canal SLAC1. En huevos de rana modificados para producir las proteínas vegetales, añadir CA4 normal aumentó la corriente iónica de SLAC1, mientras que formas mutantes de CA4 que ya no podían unirse al canal no lograron hacerlo. De manera sorprendente, un mutante de CA4 que había perdido la actividad catalítica pero seguía uniéndose a SLAC1 aún pudo aumentar la corriente. En las células guardianas de hojas de Arabidopsis, la CA4 normal restauró un fuerte aumento de la actividad de SLAC1 dependiente del dióxido de carbono. En cambio, las plantas que expresaban CA4 defectuosa en la unión mostraron poco o ningún aumento en la actividad del canal incluso con dióxido de carbono elevado, a pesar de que algunos de estos mutantes aún realizaban la química normal. Esto demuestra que el contacto físico directo de CA4 con SLAC1, y no solo su capacidad para procesar dióxido de carbono, es lo que ajusta la respuesta del canal.
De canales individuales al rendimiento de la planta entera
El equipo siguió entonces las consecuencias de interrumpir la unión CA4–SLAC1 a escala de hojas y plantas enteras. En plantas donde CA4 no podía engancharse a SLAC1, los estomas se cerraban y volvían a abrir mucho más lentamente después de que los niveles de dióxido de carbono aumentaran o regresaran a la normalidad. Modelos por ordenador predijeron este comportamiento lento y sugirieron que haría a las plantas menos eficientes en el uso del agua. Experimentos bajo luz fluctuante controlada confirmaron estas predicciones: las plantas con CA4 con la unión deteriorada tenían rosales más pequeños, menor peso seco y una eficiencia en el uso del agua significativamente peor que las plantas con CA4 normal o con CA4 deficiente en catálisis pero competente en unión. Es importante destacar que su maquinaria fotosintética en sí funcionaba igual de bien, lo que indica que la penalización en el crecimiento provenía principalmente de movimientos estomáticos mal sincronizados más que de una química defectuosa de fijación de carbono.
Qué significa esto para los cultivos del futuro
En conjunto, los resultados revelan a CA4 como un socio-sensor que se adhiere al canal SLAC1 en las células guardianas y ajusta directamente su actividad en respuesta al dióxido de carbono cercano al ambiente. Esta unión utiliza un motivo estructural distinto del centro de reacción química de la enzima, demostrando que su papel regulador puede desacoplarse de su función catalítica. Al afinar la velocidad con la que los estomas se abren y cierran, esta asociación proteica ayuda a las plantas a encontrar un mejor equilibrio entre la captación de carbono y el ahorro de agua, especialmente bajo condiciones de luz natural que cambian minuto a minuto. En términos prácticos, el trabajo apunta a nuevos objetivos moleculares para la cría o la ingeniería de cultivos cuyas válvulas foliares respondan con mayor agilidad, lo que podría aumentar los rendimientos mientras se utiliza menos agua en un mundo que se calienta y se seca.
Cita: Xia, L., Alvim, J.C., Nguyen, TH. et al. A guard cell carbonic anhydrase binds and regulates SLAC1 separate from its catalytic activity. Nat Commun 17, 3911 (2026). https://doi.org/10.1038/s41467-026-70596-9
Palabras clave: estomas, detección de dióxido de carbono, células guardianas, eficiencia en el uso del agua, canales iónicos