Clear Sky Science · de
Ein Carbonanhydrase in Schließzellen bindet und reguliert SLAC1 unabhängig von seiner katalytischen Aktivität
Warum winzige Poren in Blättern wichtig sind
Jedes Blatt ist mit mikroskopisch kleinen Poren übersät, die wie einstellbare Ventile Wasser gegen Kohlendioxid tauschen. Diese Ventile, die als Spaltöffnungen bezeichnet werden, müssen sich öffnen, damit Kohlendioxid für die Photosynthese einströmen kann, dürfen aber schnell schließen, um ein Austrocknen der Pflanze zu verhindern. Wie Spaltöffnungen Kohlendioxidwerte in Echtzeit wahrnehmen und darauf reagieren, ist seit Langem ein Rätsel. Diese Studie legt ein Schlüsselteilstück dieses Mechanismus offen und zeigt, wie zwei Proteine in den Zellen, die jede Pore umgeben, zusammenarbeiten, um Wasserverlust und Wachstum fein abzustimmen.
Blattventile, die Luft und Wasser jonglieren
Spaltöffnungen bestehen aus Paaren von Schließzellen, die sich durch Bewegung von Ionen und Wasser öffnen oder schließen. Wenn die umgebende Luft oder die Luftkammern im Blatt mehr Kohlendioxid enthalten, neigen die Spaltöffnungen dazu, zu schließen. Das schützt die Pflanze davor, Wasser zu vergeuden, wenn ohnehin reichlich Kohlendioxid vorhanden ist. Ein wichtiger Weg für die Freisetzung negativer Ionen während des Schließvorgangs ist ein Membrankanalsystem namens SLAC1. Frühere Arbeiten deuteten darauf hin, dass eine Enzymfamilie, die Carbonanhydrasen, die normalerweise die Umwandlung zwischen Kohlendioxid und Bicarbonat in Wasser beschleunigen, irgendwie dazu beitrug, SLAC1 zu aktivieren. Unklar war jedoch, ob diese Enzyme lediglich als chemische Katalysatoren wirkten oder ob sie auch als direkte Steuerknöpfe für den Kanal fungieren.
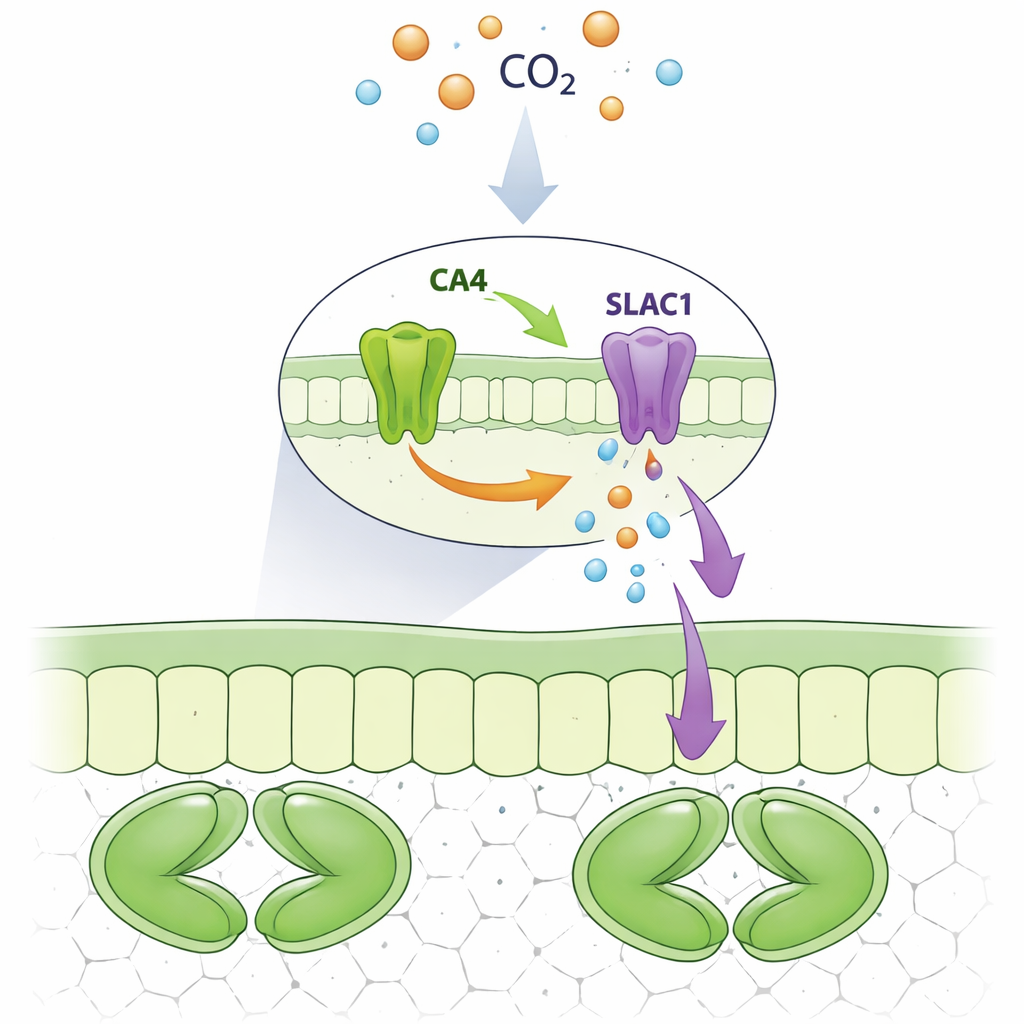
Eine Proteinpartnerschaft an der Pore
Die Forscher konzentrierten sich auf eine bestimmte Carbonanhydrase, CA4, die in Schließzellen reichlich vorkommt. Mithilfe von Hefezellen und fluoreszenzbasierter Bildgebung in Pflanzenzellen zeigten sie, dass SLAC1 einen physischen Komplex mit CA4 bildet, nicht jedoch mit eng verwandten Enzymen wie CA1 oder CA3. Beide Versionen von CA4 in Pflanzen, eine membrangebunden und eine freier in der Zellflüssigkeit, konnten an SLAC1 binden. Das Team veränderte dann systematisch einzelne Aminosäuren in CA4, um herauszufinden, welche für diese Wechselwirkung nötig sind. Sie entdeckten ein kurzes oberflächenexponiertes Stück von CA4, das um einige spezifische Reste zentriert ist und weit von seinem chemischen Reaktionszentrum entfernt liegt und das für die Bindung an SLAC1 unerlässlich ist. Mutationen in diesem Motiv zerstörten die Interaktion, während die katalytische Aktivität des Enzyms erhalten blieb.
Chemie von Kontrolle trennen
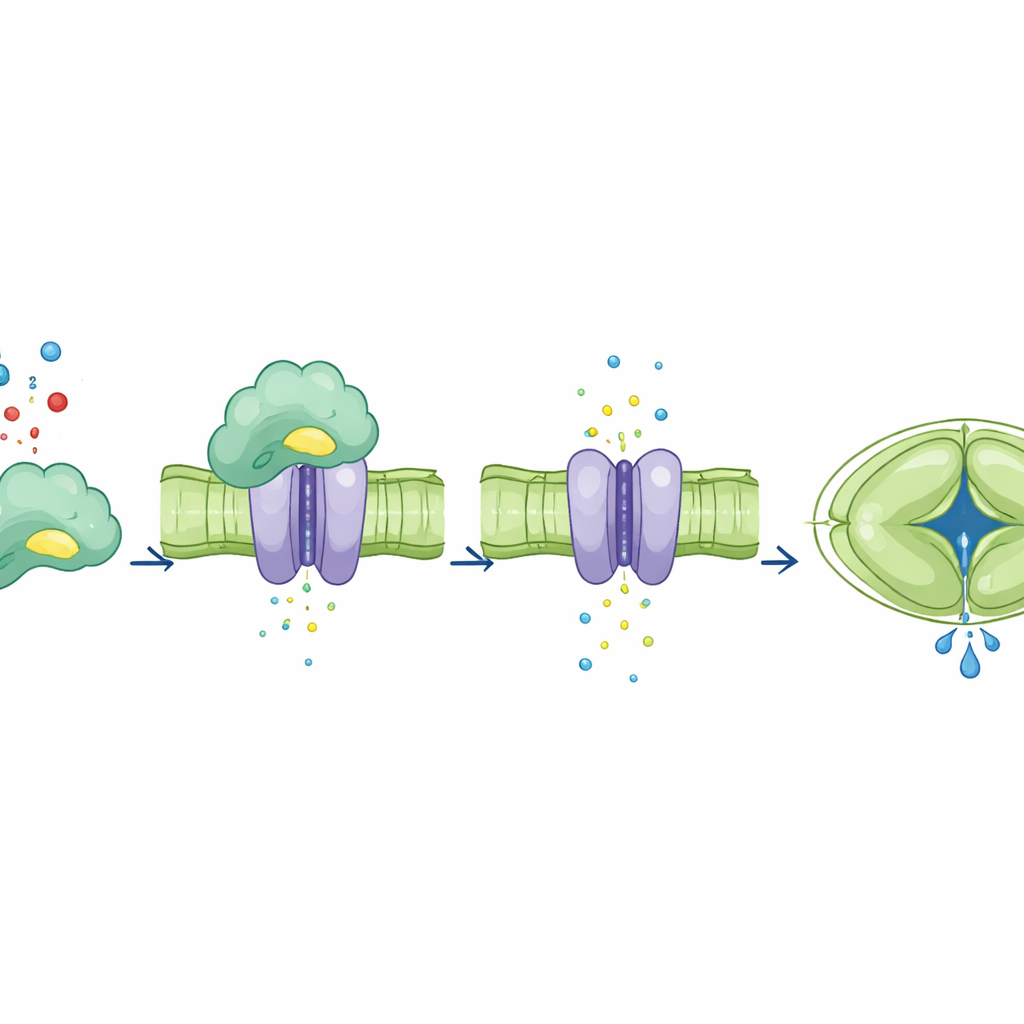
Nachdem sie CA4s Bindungsfunktion von seiner katalytischen Funktion getrennt hatten, untersuchten die Autoren, wie sich jede auf den SLAC1-Kanal auswirkt. In Froscheiern, die so verändert wurden, dass sie die Pflanzenproteine produzierten, verstärkte die Zugabe von normalem CA4 den Ionenstrom von SLAC1, während mutierte Formen von CA4, die den Kanal nicht mehr binden konnten, dies nicht vermochten. Auffällig war, dass ein CA4-Mutant, der seine katalytische Aktivität verloren hatte, aber weiterhin an SLAC1 band, den Strom trotzdem steigern konnte. In Schließzellen von Arabidopsis-Blättern stellte normales CA4 eine starke, CO2-abhängige Erhöhung der SLAC1-Aktivität wieder her. Im Gegensatz dazu zeigten Pflanzen, die bindungsdefektes CA4 exprimierten, kaum oder keine Steigerung der Kanalaktivität selbst bei erhöhtem Kohlendioxid, obwohl einige dieser Mutanten weiterhin normale chemische Aktivität zeigten. Das zeigt, dass der direkte physische Kontakt von CA4 mit SLAC1 — nicht nur seine Fähigkeit, Kohlendioxid umzuwandeln — die Reaktion des Kanals einstellt.
Von einzelnen Kanälen zur Leistung der ganzen Pflanze
Das Team verfolgte dann die Folgen einer gestörten CA4–SLAC1-Bindung auf der Ebene ganzer Blätter und Pflanzen. In Pflanzen, in denen CA4 nicht an SLAC1 andocken konnte, schlossen und öffneten sich die Spaltöffnungen nach Erhöhung oder Rückkehr des Kohlendioxidgehalts wesentlich langsamer. Computermodelle sagten dieses trägere Verhalten voraus und deuteten darauf hin, dass es Pflanzen weniger effizient im Wasserverbrauch machen würde. Experimente unter kontrollierten, schwankenden Lichtbedingungen bestätigten diese Vorhersagen: Pflanzen mit bindungsbeeinträchtigtem CA4 bildeten kleinere Rosetten, geringeres Trockengewicht und eine deutlich schlechtere Wasserverwendungseffizienz als Pflanzen mit normalem oder katalysedefizientem, aber bindungsfähigem CA4. Wichtig ist, dass die photosynthetische Maschinerie selbst ebenso gut funktionierte, was darauf hinweist, dass die Wachstumsstrafe hauptsächlich auf falsch getimte stomatale Bewegungen und nicht auf fehlerhafte Kohlenstofffixierungschemie zurückzuführen ist.
Was das für zukünftige Nutzpflanzen bedeutet
Zusammen zeigen die Ergebnisse, dass CA4 als Sensorpartner fungiert, der an den SLAC1-Kanal in Schließzellen andockt und dessen Aktivität direkt in Reaktion auf Kohlendioxid nahe der Umgebungswerte anpasst. Diese Bindung erfolgt über ein Strukturmotiv, das sich vom chemischen Reaktionszentrum des Enzyms unterscheidet, und beweist, dass seine regulatorische Rolle von seiner katalytischen Rolle entkoppelt werden kann. Indem sie die Geschwindigkeit des Öffnens und Schließens der Spaltöffnungen schärft, hilft diese Proteinpartnerschaft Pflanzen, ein besseres Gleichgewicht zwischen Kohlenstoffaufnahme und Wassersparen zu finden, besonders unter natürlichen Lichtbedingungen, die von Minute zu Minute wechseln. Praktisch gesehen weist die Arbeit auf neue molekulare Ziele für Züchtung oder gentechnische Optimierung von Kulturen hin, deren Blattventile reaktionsschneller sind — was potenziell Erträge steigern und den Wasserverbrauch in einer sich erwärmenden, austrocknenden Welt reduzieren könnte.
Zitation: Xia, L., Alvim, J.C., Nguyen, TH. et al. A guard cell carbonic anhydrase binds and regulates SLAC1 separate from its catalytic activity. Nat Commun 17, 3911 (2026). https://doi.org/10.1038/s41467-026-70596-9
Schlüsselwörter: Spaltöffnungen, CO2-Sinneswahrnehmung, Schließzellen, Wasserverwendungseffizienz, Ionenkanäle