Clear Sky Science · pl
Mechanizm ko‑transkrypcyjnego „cap snatchingu” przez polimerazę wirusa grypy
Jak wirusy grypy kradną znaczniki z naszych komórkowych wiadomości
Każdej zimy wirusy grypy rozprzestrzeniają się po populacji, a ich powodzenie zależy od zaskakująco precyzyjnego skoku, który rozgrywa się głęboko w jądrach naszych komórek. Aby tworzyć własne komunikaty genetyczne, wirusy grypy nie potrafią wytworzyć ochronnej „czapeczki” (cap), którą komórki normalnie doklejają na początku swoich RNA. Zamiast tego kradną tę czapeczkę bezpośrednio z właśnie zsyntetyzowanych RNA gospodarza w manewrze zwanym „cap snatching”. Badanie to pokazuje, z molekularną szczegółowością, jak wirus grypy przyczepia się do ludzkiego aparatu transkrypcyjnego i odcina te czapeczki, by napędzać własną replikację.
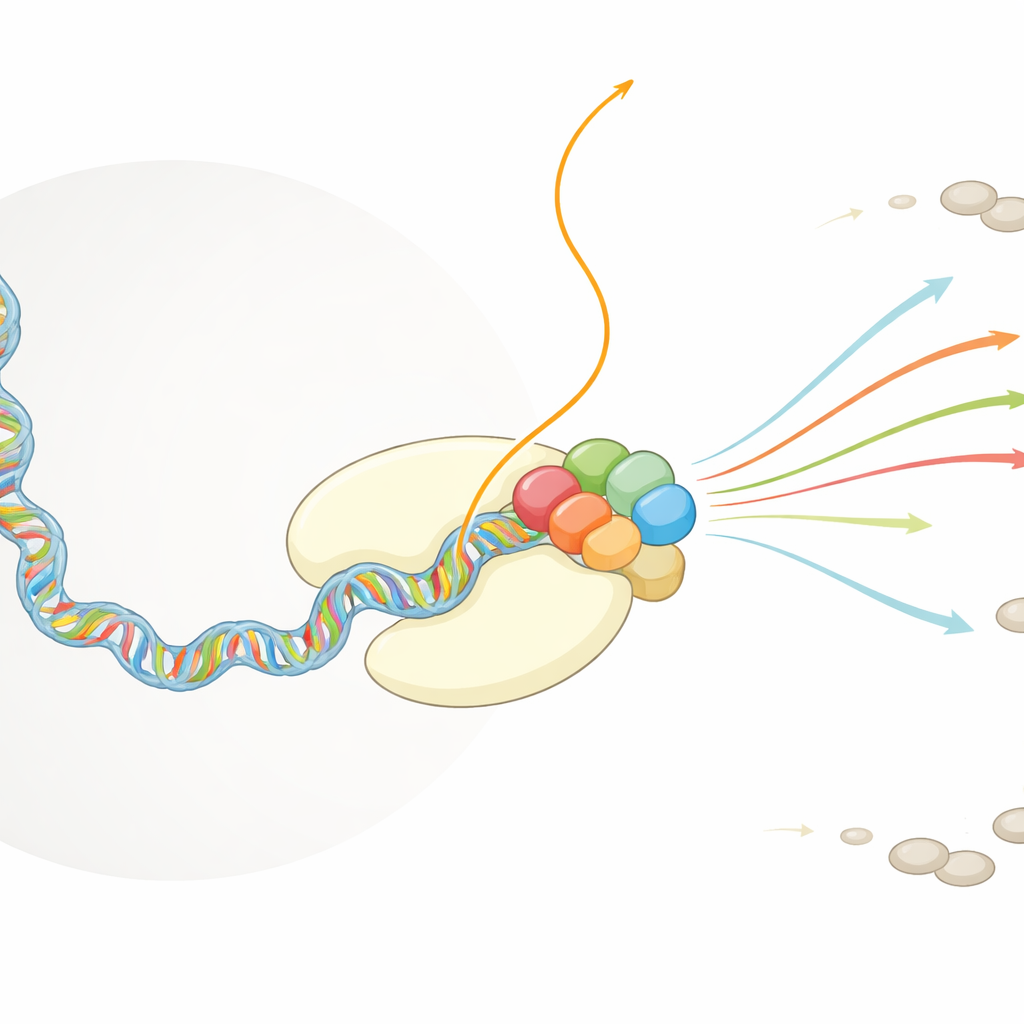
Kopiarka komórkowa i pasożyt wirusowy
Ludzkie komórki polegają na ogromnym kompleksie białkowym, polimerazie RNA II, który odczytuje DNA i produkuje kopie RNA, które potem zostaną przetłumaczone na białka. Gdy RNA wychodzi z tej maszyny, inne czynniki komórkowe szybko ozdabiają jego początek ochronną czapeczką, która stabilizuje molekułę i zapewnia jej eksport z jądra oraz tłumaczenie. Wirus grypy działa w tej samej przestrzeni jądrowej. Jego własny kompleks polimerazy, zwany FluPol, nie potrafi wytwarzać capów, lecz zależy całkowicie od spójnych z capem fragmentów RNA, aby rozpocząć przepisywanie genomu wirusowego na mRNA. Wcześniejsze prace wykazały, że FluPol wiąże się z polimerazą RNA II, ale nie było wiadomo, w jaki sposób to połączenie ustawia FluPol, by chwycić i przeciąć RNA gospodarza.
Uchwycenie partnerstwa wirusa i gospodarza w działaniu
Aby rozpracować ten proces, badacze odtworzyli w probówce kluczową część ludzkiego aparatu transkrypcyjnego: polimerazę RNA II związana z DNA, produkującą capowane RNA, oraz czynnik elongacyjny DSIF, który chwyta powstające RNA. Następnie dodali FluPol i zastosowali krioelektronową mikroskopię, by zwizualizować powstały kompleks w niemal atomowej rozdzielczości, zarówno przed, jak i po przecięciu przez wirusa. Równoległe testy biochemiczne mierzyły, jak efektywnie FluPol przecina RNA gospodarza w różnych warunkach, na przykład z obecnymi lub brakującymi chemicznymi modyfikacjami ogona polimerazy II albo przy udziale DSIF.
Gdzie i jak dokonywane jest cięcie wirusa
Struktury pokazują, że FluPol osiada przy boku polimerazy RNA II dokładnie tam, gdzie wychodzi nowe RNA. Jedna część FluPol rozpoznaje zmodyfikowany ogon polimerazy II, gdy niesie specyficzne grupy fosforanowe — znak bardzo wczesnej transkrypcji. Inna część, region PB2 „wiązania capa”, wciska się w rowek na powierzchni polimerazy II tuż pod kanałem wyjścia RNA i zaciska się na capie na początku RNA. W międzyczasie region PA, funkcjonujący jako endonukleaza FluPol, nawiązuje kontakt z DSIF, który obrócił się ze swojej zwykłej pozycji, by zrobić miejsce i poprowadzić RNA w stronę miejsca cięcia wirusa. Te kontakty stabilizują kompleks tak, że endonukleaza może odciąć RNA gospodarza w odległości około 10–15 nukleotydów od capa, generując krótki, capowany fragment, którego wirus może użyć.
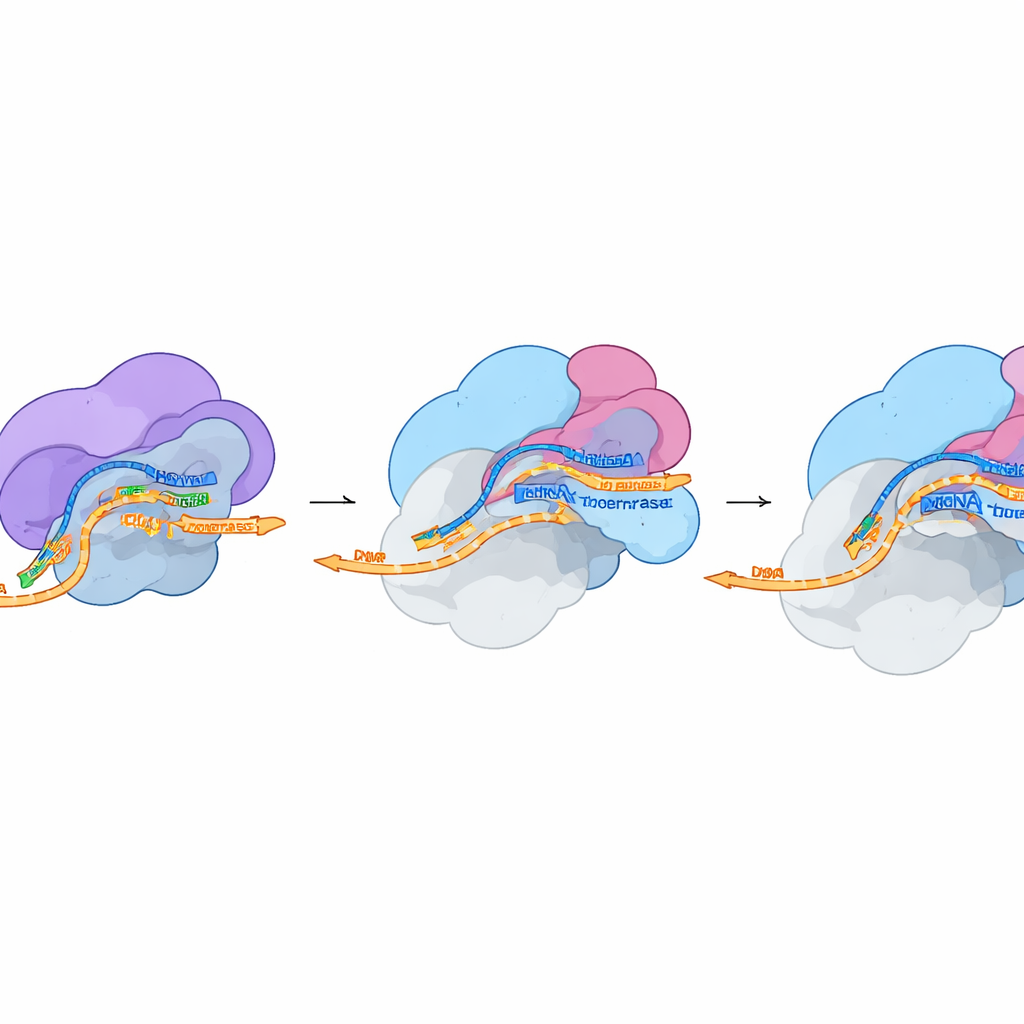
Od skradzionego fragmentu do wirusowej wiadomości
W drugim ujęciu strukturalnym wykonanym w warunkach umożliwiających cięcie, badacze stwierdzili, że FluPol pozostaje przyłączony do aparatu gospodarza nawet po przecięciu RNA. Cap pozostaje mocno utrzymany w kieszeni PB2, ale świeżo odsłonięty koniec skradzionego fragmentu obraca się w stronę centrum aktywnego FluPol, gdzie rozpoczyna się przepisywanie wirusowego RNA. Ogólny kształt FluPol w tym stanie odpowiada wcześniej opisanej formie „pre‑inicjacyjnej”, gotowej do rozpoczęcia syntezy. Sugeruje to, że do przejścia od cap snatchingu do pełnej transkrypcji wirusowej potrzebne są jedynie minimalne zmiany konformacyjne. Doświadczenia w komórkach, w których pojedyncze punkty kontaktowe między FluPol, DSIF i polimerazą II zostały zmutowane, wykazały, że zaburzenie tych subtelnych interfejsów ostro redukuje aktywność polimerazy wirusowej i w niektórych przypadkach wpływa negatywnie na kondycję wirusa.
Dlaczego te odkrycia są istotne
Praca ujawnia, że wirus grypy nie chwyta capów z dowolnego RNA; celuje w bardzo wczesny etap ludzkiej transkrypcji, gdy polimeraza II, jej zmodyfikowany ogon, DSIF i świeżo powstały cap spotykają się razem. FluPol dokuje na tej złożonej platformie, wykorzystuje DSIF do przedstawienia RNA swojemu miejscu cięcia, odcina capowany fragment, a następnie podaje go bezpośrednio do własnego aparatu kopiującego, by rozpocząć produkcję wirusowego mRNA. Dla laika kluczowy wniosek jest taki, że sukces grypy zależy od precyzyjnego fizycznego objęcia między białkami wirusa i gospodarza. Choć takie małe, płaskie powierzchnie kontaktu są trudne do zaatakowania lekami, te mapy na poziomie atomowym dokładnie wskazują, gdzie potencjalny przyszły środek przeciwwirusowy mógłby się wcisnąć, by przerwać kradzież i osłabić infekcję grypową.
Cytowanie: Rotsch, A.H., Li, D., Dupont, M. et al. Mechanism of co-transcriptional cap snatching by influenza polymerase. Nature 652, 1281–1288 (2026). https://doi.org/10.1038/s41586-026-10189-0
Słowa kluczowe: wirus grypy, cap snatching, polimeraza RNA II, transkrypcja wirusowa, struktura cryo‑EM