Clear Sky Science · de
Mechanismus des kofraktionellen „cap snatching“ durch die Influenzapolymerase
Wie Grippeviren die Kopftags unserer Zellen stehlen
Jeden Winter verbreiten sich Influenzaviren in der menschlichen Bevölkerung, doch ihr Erfolg beruht auf einem überraschend filigranen Diebstahl, der tief im Zellkern abläuft. Um ihre eigenen genetischen Botschaften zu erzeugen, können Grippeviren die schützende "Kappe" am 5'-Ende der RNA, die Zellen normalerweise anbringen, nicht selbst herstellen. Stattdessen stehlen sie diese Kappe direkt von frisch synthetisierten zellulären RNAs in einem Manöver, das "cap snatching" genannt wird. Diese Studie zeigt auf molekularer Ebene, wie das Influenzavirus die menschliche Transkriptionsmaschinerie anbindet und diese Kappen abschneidet, um seine eigene Replikation anzutreiben.
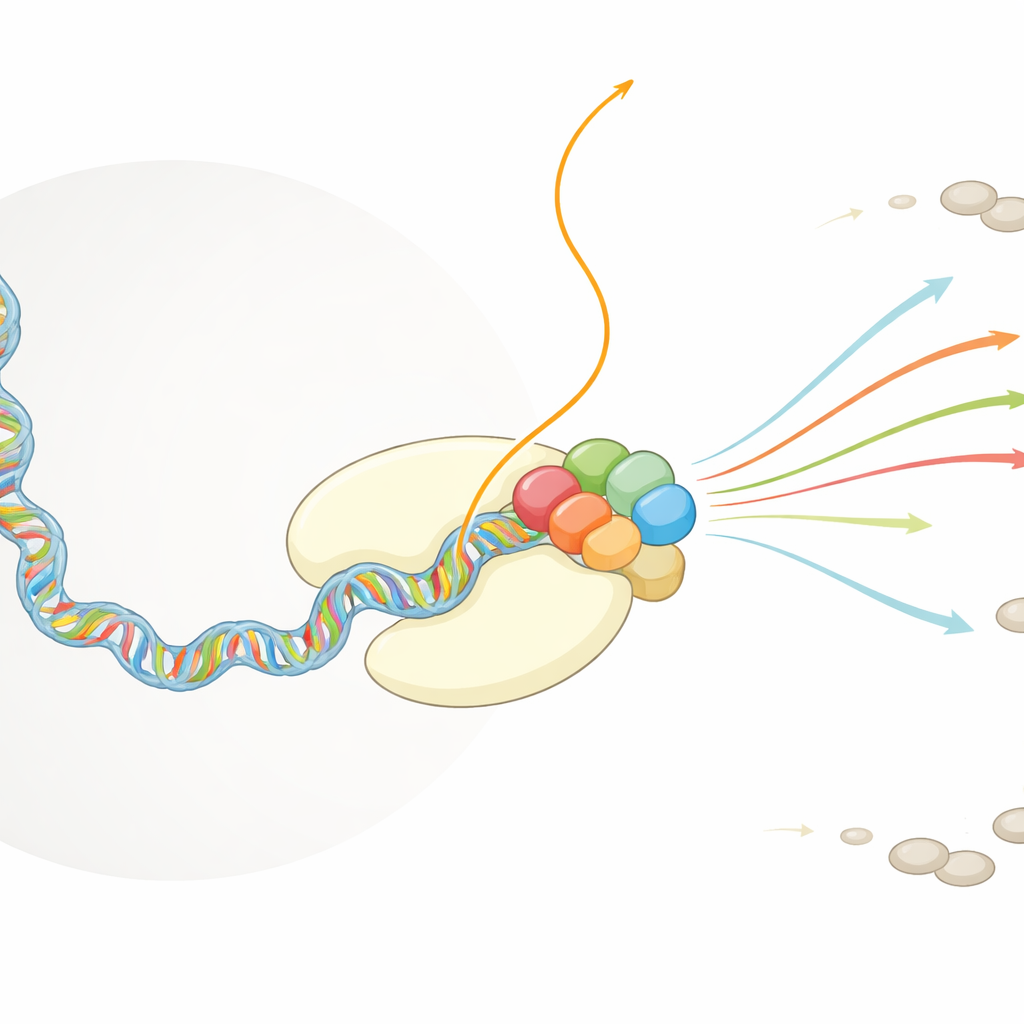
Die Kopiermaschine der Zelle und der virale Mitfahrer
Humanen Zellen sind auf einen riesigen Proteinkomplex, die RNA-Polymerase II, angewiesen, um DNA abzulesen und RNA-Kopien zu erzeugen, die später in Proteine übersetzt werden. Sobald eine RNA aus dieser Maschine herauskommt, schmücken andere zelluläre Faktoren ihr 5'-Ende rasch mit einer schützenden Kappstruktur, die das Molekül stabilisiert und dafür sorgt, dass es aus dem Kern exportiert und translatiert wird. Das Influenzavirus operiert im selben nukleären Raum. Sein eigener Polymerasekomplex, FluPol genannt, kann keine Kappen herstellen, ist aber völlig abhängig von gekappten RNAfragmenten, um die virale Genome in mRNAs zu überführen. Frühere Arbeiten zeigten, dass FluPol an die RNA-Polymerase II bindet, aber wie diese Wechselwirkung FluPol so positioniert, dass es zelluläre RNA greifen und schneiden kann, war unbekannt.
Die Flu–Wirt-Partnerschaft in Aktion einfangen
Um diesen Prozess zu zerlegen, rekonstruierten die Forscher einen Schlüsselteil der menschlichen Transkriptionsmaschinerie im Reagenzglas: RNA-Polymerase II auf DNA, die eine gekappte RNA produziert, begleitet von einem Elongationsfaktor namens DSIF, der die entstehende RNA umgreift. Anschließend fügten sie FluPol hinzu und verwendeten kryo-elektronenmikroskopie, um das kombinierte Komplex bei annähernd atomarer Auflösung sowohl vor als auch nach dem viralen Schnitt zu visualisieren. Parallele biochemische Assays maßen, wie effizient FluPol zelluläre RNA unter verschiedenen Bedingungen spalten konnte, etwa mit oder ohne chemische Markierungen am Schwanz der RNA-Polymerase II oder bei Anwesenheit von DSIF.
Wo und wie der virale Schnitt erfolgt
Die Strukturen zeigen FluPol, das sich an die Seite der RNA-Polymerase II schmiegt, genau dort, wo die neue RNA austritt. Ein Teil von FluPol erkennt den modifizierten Schwanz der Polymerase II, wenn dieser spezifische Phosphatgruppen trägt — ein Kennzeichen sehr früher Transkriptionsphasen. Ein anderer Abschnitt, die PB2-"cap-binding"-Region, dringt in eine Rinne auf der Oberfläche der Polymerase II direkt unterhalb des RNA-Austrittskanals ein und klemmt die Kappe am 5'-Ende der RNA fest. Unterdessen kontaktiert die PA-"Endonuclease"-Region von FluPol DSIF, das sich von seiner üblichen Position weggedreht hat, um Platz zu machen und die RNA zum viralen Schnittplatz zu führen. Zusammen stabilisieren diese Kontakte den Komplex so, dass die Endonuclease die Wirts-RNA etwa 10–15 Nukleotide von der Kappe entfernt aufschneiden kann und ein kurzes, gekapptes Fragment entsteht, das das Virus nutzen kann.
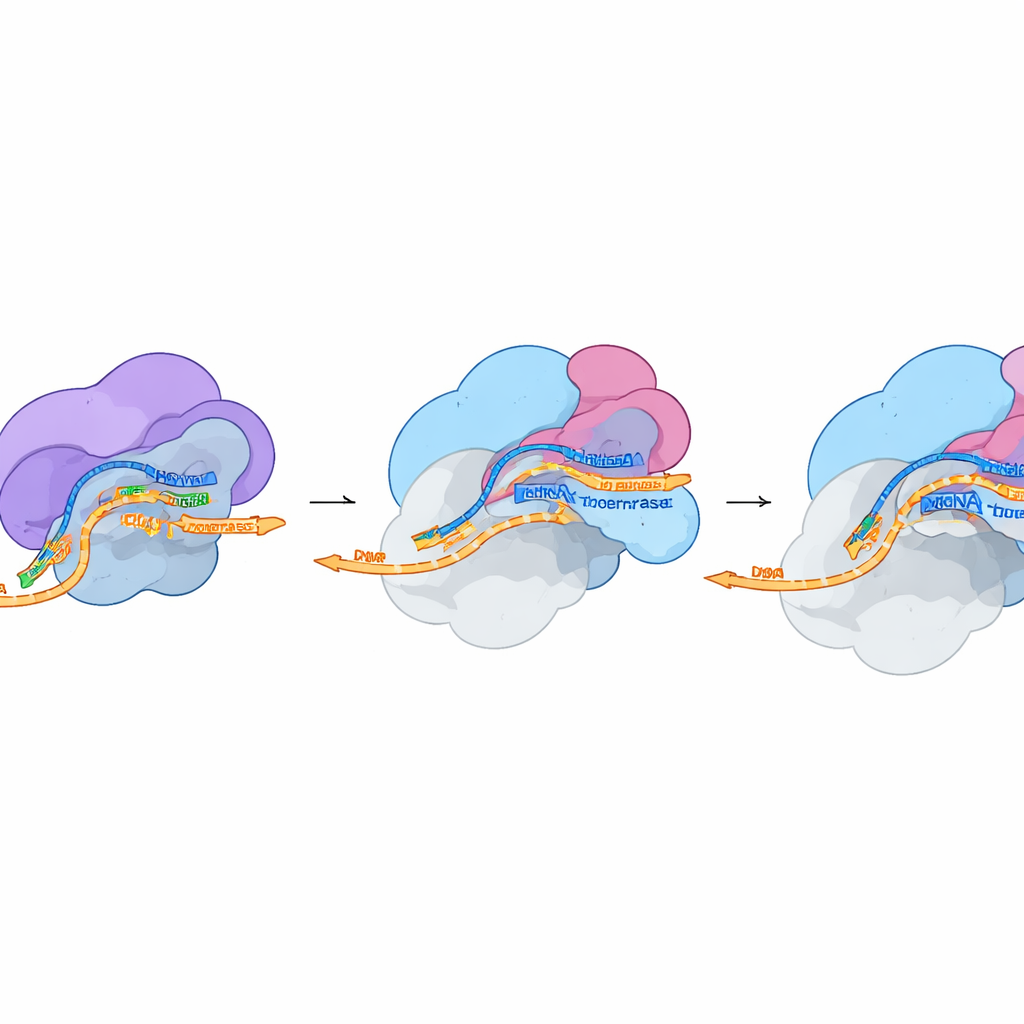
Vom gestohlenen Fragment zur viralen Botschaft
In einer zweiten strukturellen Momentaufnahme unter schneidefähigen Bedingungen stellten die Forscher fest, dass FluPol auch nach der RNA-Spaltung an der Wirtsmaschinerie befestigt bleibt. Die Kappe sitzt weiterhin fest in der PB2-Tasche, aber das frisch freigelegte Ende des gestohlenen Fragments schwenkt in Richtung des aktiven Zentrums von FluPol, wo die Kopie der viralen RNA beginnt. Die Gesamtform von FluPol in diesem Zustand entspricht weitgehend einer zuvor beschriebenen "Pre-Initiation"-Form, die bereit ist, die Synthese zu starten. Das deutet darauf hin, dass nur minimale Konformationsänderungen nötig sind, um vom cap snatching zur vollständigen viralen Transkription überzugehen. Zellbasierte Experimente, in denen einzelne Kontaktpunkte zwischen FluPol, DSIF und der Polymerase II mutiert wurden, zeigten, dass die Störung dieser kleinen Schnittstellen die Aktivität der viralen Polymerase stark reduziert und in einigen Fällen die gesamte Virusfitness mindert.
Warum diese Erkenntnisse wichtig sind
Diese Arbeit zeigt, dass die Influenza nicht einfach Kappen von beliebigen RNAs greift; sie zielt auf eine sehr frühe Phase der menschlichen Transkription ab, in der Polymerase II, ihr modifizierter Schwanz, DSIF und eine neu abgeschlossene Kappe zusammenkommen. FluPol dockt an dieser zusammengesetzten Plattform an, nutzt DSIF, um die RNA dem Schnittplatz zu präsentieren, trennt ein gekapptes Fragment ab und schiebt dieses Fragment direkt in seine eigene Kopiermaschinerie, um die Produktion viraler mRNAs zu starten. Für den allgemein Interessierten lautet die zentrale Erkenntnis: Der Erfolg der Grippe hängt von einer präzisen physischen Umarmung zwischen viralen und zellulären Proteinen ab. Obwohl solche kleinen, flachen Kontaktflächen schwierige Wirkstoffziele sind, zeigen diese atomaren Karten nun genau, wo ein zukünftiges antivirales Mittel eingreifen könnte, um den Diebstahl zu stören und die Influenzainfektion abzuschwächen.
Zitation: Rotsch, A.H., Li, D., Dupont, M. et al. Mechanism of co-transcriptional cap snatching by influenza polymerase. Nature 652, 1281–1288 (2026). https://doi.org/10.1038/s41586-026-10189-0
Schlüsselwörter: Influenzavirus, cap snatching, RNA-Polymerase II, virale Transkription, cryo-EM-Struktur