Clear Sky Science · it
Meccanismo del "cap snatching" co‑trascrizionale da parte della polimerasi dell’influenza
Come i virus dell’influenza rubano le etichette dei nostri messaggi cellulari
Ogni inverno i virus influenzali si propagano nelle popolazioni umane, ma il loro successo dipende da un furto sorprendentemente delicato che avviene nel profondo del nucleo delle nostre cellule. Per costruire i propri messaggi genetici, i virus dell’influenza non possono sintetizzare il “cap” protettivo che le cellule aggiungono normalmente all’estremità 5' degli RNA. Invece, lo sottraggono direttamente dai nostri RNA neoformati in una manovra chiamata “cap snatching”. Questo studio rivela, a livello molecolare, come il virus dell’influenza si aggancia alla macchina trascrizionale umana e recide questi cap per alimentare la propria replicazione.
La fotocopiatrice della cellula e il passeggero virale
Le cellule umane si affidano a un gigantesco complesso proteico, l’RNA polimerasi II, per leggere il DNA e produrre copie di RNA che verranno poi tradotte in proteine. Non appena un RNA emerge da questa macchina, altri fattori cellulari decorano rapidamente la sua estremità con una struttura di cap protettiva che stabilizza la molecola e ne garantisce l’esportazione dal nucleo e la traduzione. Il virus dell’influenza opera nello stesso spazio nucleare. Il suo complesso di polimerasi, chiamato FluPol, non è in grado di sintetizzare i cap ma dipende in modo assoluto da frammenti di RNA incappati per iniziare a trascrivere il genoma virale in mRNA. Lavori precedenti avevano mostrato che FluPol si lega all’RNA polimerasi II, ma restava ignoto come questa interazione posizionasse FluPol per afferrare e tagliare l’RNA ospite.
Cogliere in azione la partnership influenza‑ospite
Per analizzare questo processo, i ricercatori hanno ricostituito in provetta una porzione chiave dell’apparato trascrizionale umano: l’RNA polimerasi II impegnata sul DNA, che produce un RNA incappato, accompagnata da un fattore di elongazione chiamato DSIF che afferra l’RNA emergente. Hanno quindi aggiunto FluPol e usato la microscopia crioelettronica per visualizzare il complesso combinato a risoluzione prossima a quella atomica, sia prima sia dopo il taglio virale. Saggi biochimici paralleli hanno misurato quanto efficacemente FluPol potesse clivare l’RNA ospite in diverse condizioni, ad esempio con o senza marchi chimici sulla coda dell’RNA polimerasi II o con DSIF presente.
Dove e come avviene il taglio virale
Le strutture mostrano FluPol adagiata contro il fianco dell’RNA polimerasi II esattamente nel punto in cui esce il nuovo RNA. Una porzione di FluPol riconosce la coda modificata della polimerasi II quando porta gruppi fosfato specifici—un segno caratteristico delle fasi molto precoci della trascrizione. Un’altra parte, la regione PB2 detta “cap‑binding”, si infila in una scanalatura sulla superficie della polimerasi II subito sotto il canale di uscita dell’RNA e si aggrappa al cap dell’estremità dell’RNA. Nel frattempo, la regione PA con attività endonucleasica di FluPol entra in contatto con DSIF, che ruota dalla sua posizione abituale per fare spazio e contribuire a guidare l’RNA verso il sito di taglio virale. Questi contatti stabilizzano il complesso in modo che l’endonucleasi possa sezionare l’RNA dell’ospite a circa 10–15 nucleotidi dal cap, generando un breve frammento incappato che il virus può utilizzare.
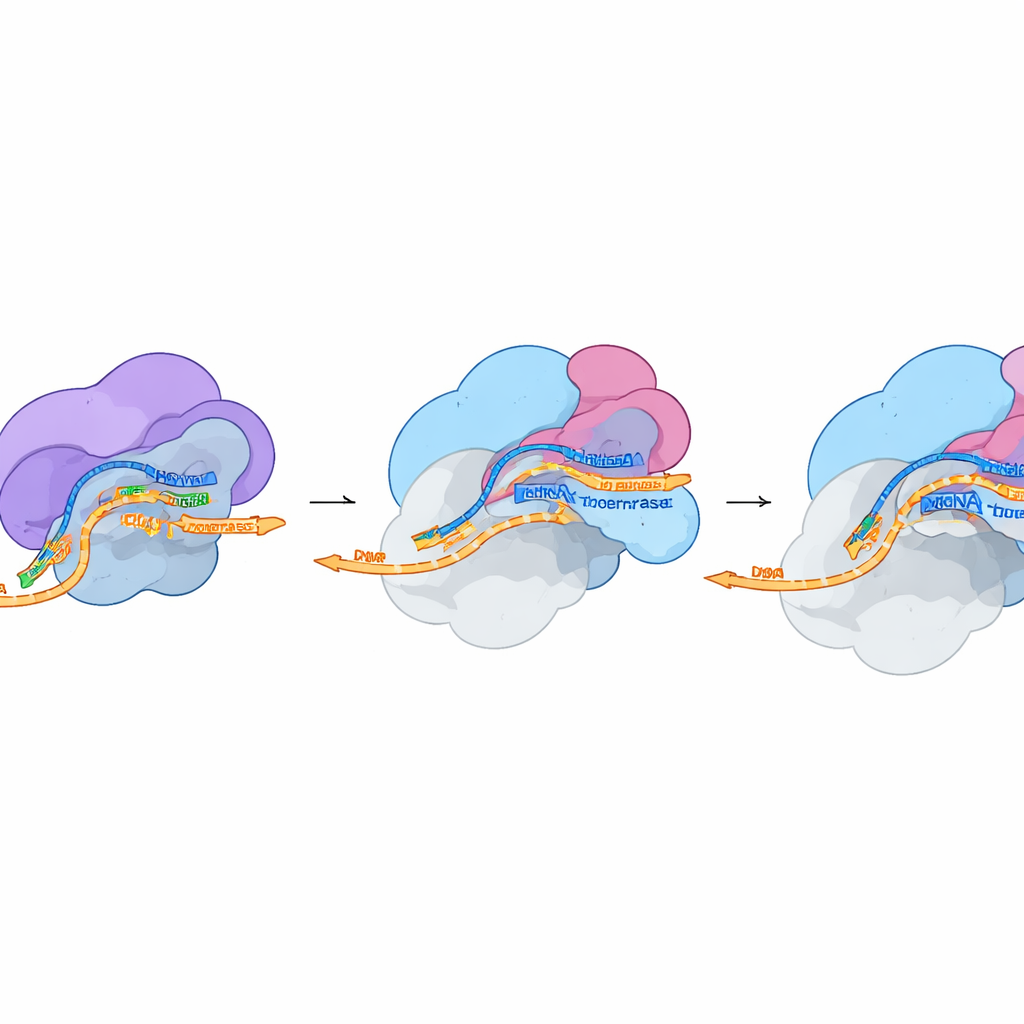
Dal frammento rubato al messaggio virale
In un secondo istantaneo strutturale ottenuto in condizioni che permettono il taglio, i ricercatori hanno osservato che FluPol rimane attaccata alla macchina ospite anche dopo che l’RNA è stato clivato. Il cap resta saldamente trattenuto nella tasca di PB2, ma l’estremità appena esposta del frammento sottratto si sposta verso il centro attivo di FluPol, dove ha inizio la copia dell’RNA virale. La conformazione complessiva di FluPol in questo stato corrisponde da vicino a una forma di “pre‑iniziazione” descritta in studi precedenti, pronta ad avviare la sintesi. Ciò suggerisce che siano necessari solo cambiamenti conformazionali minimi per passare dal cap snatching alla trascrizione virale vera e propria. Esperimenti cellulari, in cui punti di contatto specifici tra FluPol, DSIF e la polimerasi II sono stati mutati, hanno mostrato che la perturbazione di queste interfacce riduce nettamente l’attività della polimerasi virale e, in alcuni casi, la fitness complessiva del virus.
Perché questi risultati sono importanti
Questo lavoro dimostra che l’influenza non sottrae cap da qualunque RNA; mira a una fase molto precoce della trascrizione umana in cui polimerasi II, la sua coda modificata, DSIF e un cap appena formato si trovano insieme. FluPol si aggancia a questa piattaforma composita, usa DSIF per presentare l’RNA al suo sito di taglio, recide un frammento incappato e quindi inserisce quel frammento direttamente nella propria macchina di copia per avviare la produzione di mRNA virale. Per un osservatore non specialistico, la conclusione chiave è che il successo dell’influenza dipende da un abbraccio fisico preciso tra proteine virali e ospiti. Sebbene superfici di contatto così piccole e piatte siano sfidanti come bersagli farmacologici, queste mappe a livello atomico indicano esattamente dove un futuro antivirale potrebbe inserirsi per interrompere il furto e attenuare l’infezione da influenza.
Citazione: Rotsch, A.H., Li, D., Dupont, M. et al. Mechanism of co-transcriptional cap snatching by influenza polymerase. Nature 652, 1281–1288 (2026). https://doi.org/10.1038/s41586-026-10189-0
Parole chiave: virus dell’influenza, cap snatching, RNA polimerasi II, trascrizione virale, struttura cryo-EM