Clear Sky Science · pl
Petlowe pętle pore interaktywne dwóch podjednostek ATPazy determinują efektywność degradacji 26S proteasomu
Jak komórkowa maszyna do recyklingu chwyta uszkodzone białka
Każda komórka musi nieustannie oczyszczać się z uszkodzonych lub niepotrzebnych już białek. Ogromna maszyna molekularna zwana proteasomem wykonuje tę pracę, wciągając białka, miażdżąc je i uwalniając fragmenty. To badanie stawia zaskakująco szczegółowe pytanie o znaczących konsekwencjach: jak zaledwie dwa drobne elementy chwytne wewnątrz proteasomu decydują, czy białko zostanie wydajnie zniszczone, czy pozwoli się wymknąć?
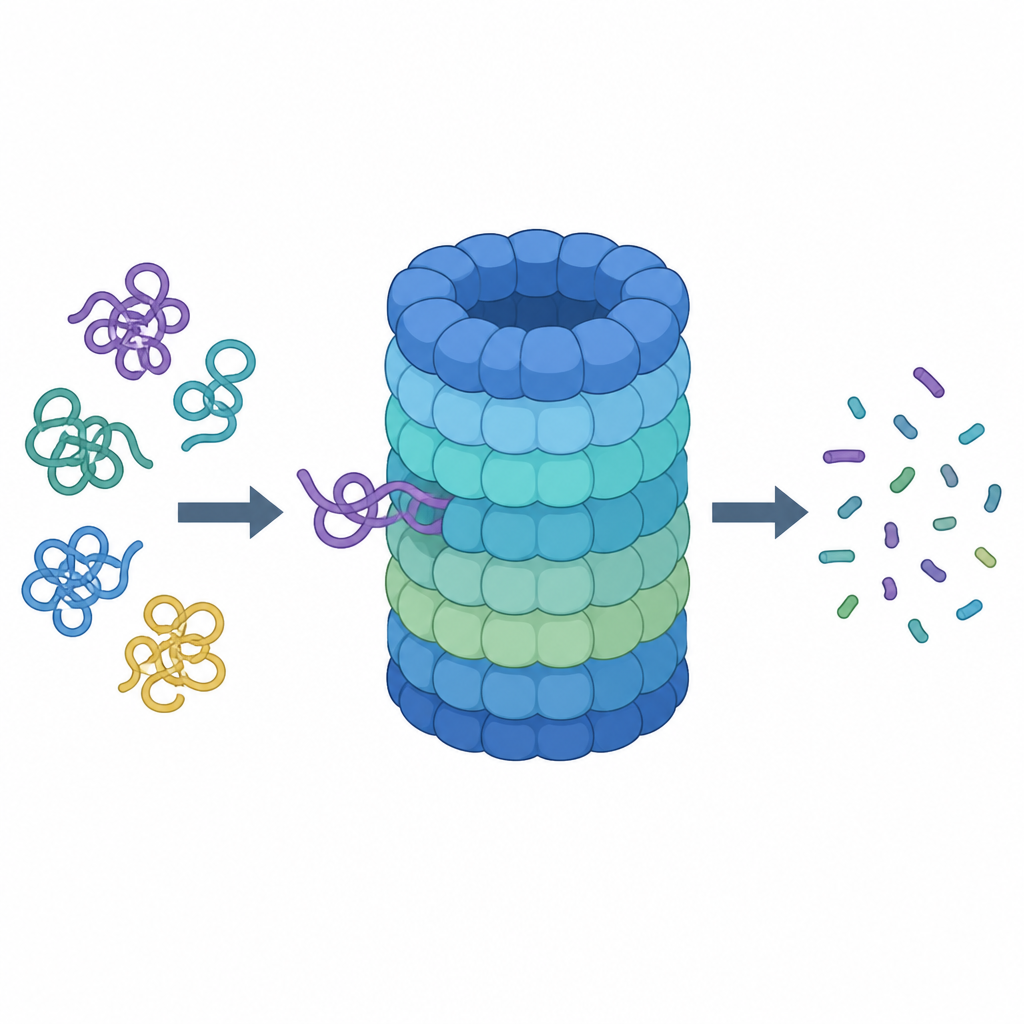
Bliższe spojrzenie na komórkową niszczarkę białek
Proteasom to maszyna w kształcie beczki złożona z wielu białkowych podjednostek. Jej centralna komora to miejsce cięcia, natomiast nasadka na górze rozpoznaje białka oznaczone do zniszczenia, usuwa ich molekularne „tagi” i wciąga je do środka. Aby zostać przyjętym jako odpad, białko musi nosić łańcuchy małej cząsteczki oznaczającej — ubikwityny — i odsłonić wiotki koniec, który można chwycić i pociągnąć. Na wejściu do nasadki znajduje się pierścień sześciu jednostek motorowych, które spalają paliwo chemiczne (ATP) i używają palcowatych pętli wewnątrz wąskiego kanału do chwytania i ciągnięcia końca łańcucha białkowego.
Dlaczego kilka pętelek chwytających ma tak duże znaczenie
Każda z sześciu jednostek motorowych ma parę pętli pore, które sięgają do centralnego tunelu jak haki i stykają się z przechodzącym łańcuchem białkowym. Wcześniejsze strukturalne migawki pokazały te haki ułożone jak spiralne schody wokół tunelu, chwytające i ciągnące na zmianę. Ale nie wszystkie haki wydawały się równe. Aby to sprawdzić, badacze osłabili jeden kluczowy aminokwas w pętli pore-1 każdej jednostki motorowej w proteasomach drożdży, a potem zmierzyli, jak dobrze maszyna zużywa ATP, zmienia kształt i rozkłada białka modelowe.
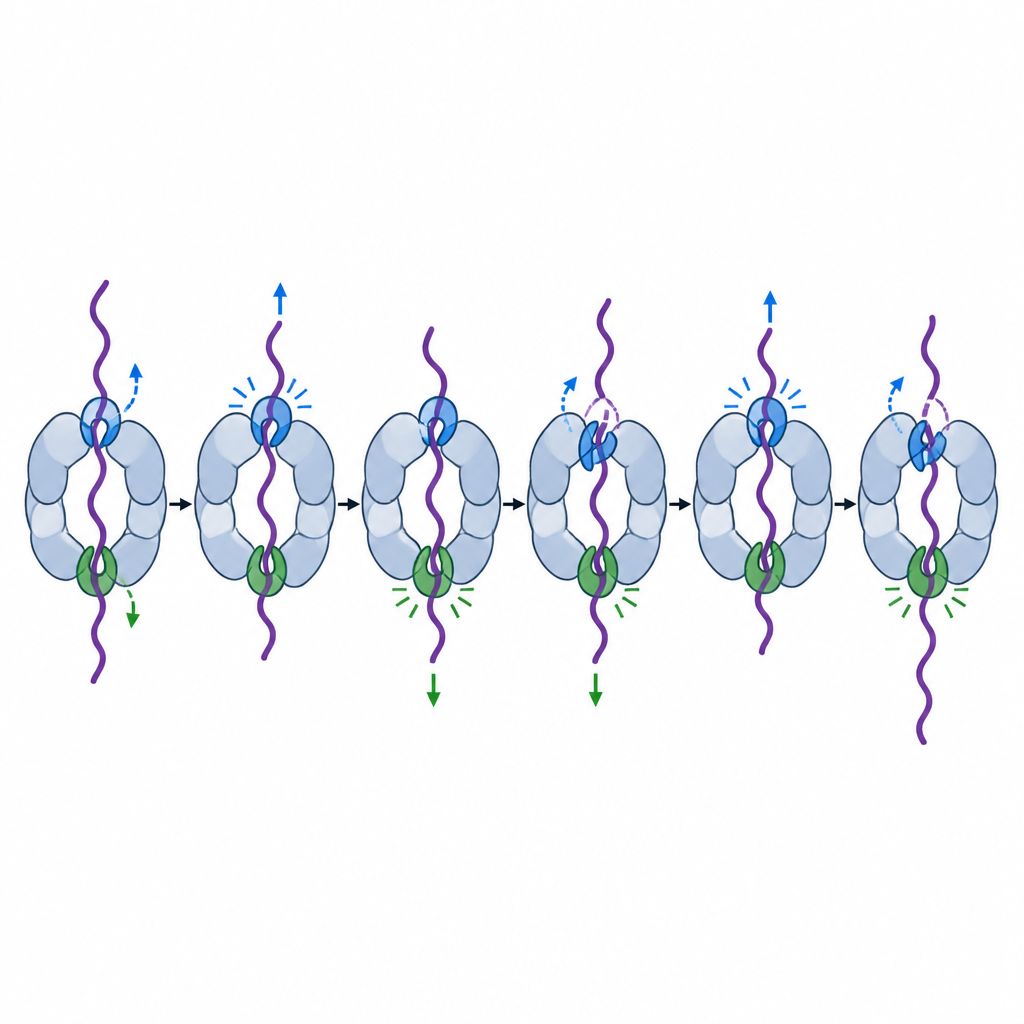
Dwa specjalne haki, które kierują uchwyceniem i rozwijaniem
Łącząc testy biochemii makroskopowej, śledzenie fluorescencyjne pojedynczych cząsteczek i wysokorozdzielczą kryoelektronową mikroskopię, zespół odkrył, że pętle pore w dwóch konkretnych jednostkach motorowych, nazwanych Rpt6 i Rpt4, pełnią szczególnie ważne, lecz odmienne role. Gdy pętla Rpt6 została osłabiona, proteasom zużywał ATP nawet bez ładunku i spędzał więcej czasu w „pozycji przetwarzania”, która częściowo blokuje wejście. Ten mutant często nie potrafił pewnie uchwycić nadchodzących końców białkowych, a nawet po rozpoczęciu pracy nad stabilnym białkiem, ciągle się ślizgał, zajmował więcej czasu lub ostatecznie pozwalał substratowi uciec. Obrazy z kriomikroskopii wyjaśniły dlaczego: w stanie spoczynkowym pętla Rpt6 jest schowana w kształcie helisy i utrzymywana przez niezwykłe kontakty z sąsiednią podjednostką, najwyraźniej blokując maszynę w cichej, gotowej do zaangażowania konfiguracji do czasu przybycia białka.
Utrzymanie mocnego uchwytu podczas trudnych zadań
Pętla Rpt4 miała inną specjalizację. Proteasomy z osłabionym hakiem Rpt4 wciąż potrafiły rozpoznać i związać oznakowane białka, ale gdy próbowały rozwinąć szczególnie odporną domenę białkową, często się ślizgały i zamiast przeciągnąć ją całkowicie, uwalniały ją. Ślady pojedynczych cząsteczek pokazywały powtarzające się próby rozwinięcia tego samego białka, przerywane krótkimi powrotami do stanu zrelaksowanego, jakby maszyna chwilowo traciła uchwyt i musiała zaczynać od nowa. Porównania strukturalne z wcześniejszymi ujęciami spiralnych stopni sugerują, że Rpt4 często znajduje się w kluczowej pozycji „szwu” tuż przed skurczem mocy, co czyni go pierwszym hakiem, który zaciska się podczas silnego pociągnięcia.
Asymetryczny silnik nastawiony na niezawodność
Ogólnie wyniki przedstawiają motor proteasomu jako asymetryczny silnik, a nie idealnie równy, sześcioczęściowy rotor. Rpt6 pomaga wyczuć, kiedy białko jest na miejscu i wywołać przejście ze stanu oczekiwania do stanu pracy, a także ponownie zaangażować łańcuch po okazjonalnych poślizgach. Z kolei Rpt4 dostarcza dużej części siły ciągnięcia potrzebnej do rozwinięcia opornych białek bez ich upuszczenia. Przypisując te odrębne zadania różnym hakom wokół pierścienia, proteasom może zarówno unikać marnowania energii, jak i zapewnić, że gdy białko zostanie wybrane do zniszczenia, zwykle zostanie rozłożone całkowicie, a nie częściowo uwolnione.
Cytowanie: López-Alfonzo, E., Saurabh, A., Zarafshan, S. et al. Substrate-interacting pore loops of two ATPase subunits determine the degradation efficiency of the 26S proteasome. Nat Commun 17, 4473 (2026). https://doi.org/10.1038/s41467-026-70426-y
Słowa kluczowe: proteasom, degradacja białek, motor ATPazy, układ ubikwityny, FRET pojedynczej cząsteczki