Clear Sky Science · es
Bucles de poro que interactúan con el sustrato en dos subunidades ATPasa determinan la eficiencia de degradación del proteasoma 26S
Cómo la máquina de reciclaje celular agarra las proteínas dañadas
Cada célula debe limpiar constantemente las proteínas dañadas o que ya no necesita. Una gigantesca máquina molecular llamada proteasoma realiza esta tarea tirando de las proteínas, triturándolas y liberando los fragmentos. Este estudio plantea una pregunta sorprendentemente concreta con grandes consecuencias: ¿cómo deciden apenas dos diminutas piezas de agarre dentro del proteasoma si una proteína será destruida eficientemente o podrá escapar?
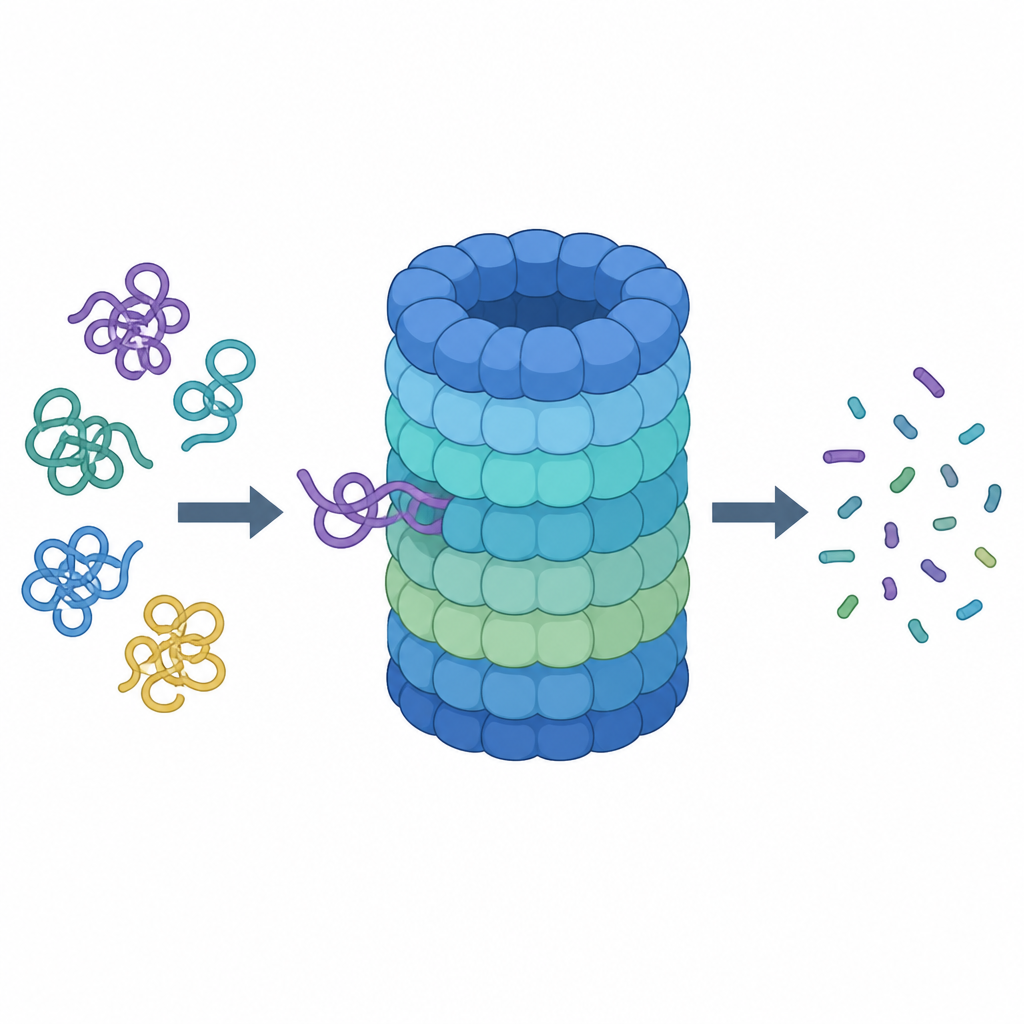
Una mirada más cercana al triturador de proteínas celular
El proteasoma es una máquina con forma de barril formada por muchas piezas proteicas. Su barril central es la cámara de corte, mientras que una tapa en la parte superior reconoce proteínas marcadas para la destrucción, elimina sus “etiquetas” moleculares y las introduce en su interior. Para ser aceptada como desecho, una proteína debe llevar cadenas de una pequeña molécula etiqueta llamada ubiquitina y exponer una cola flexible que pueda ser agarrada y tirada. En la entrada de la tapa se encuentra un anillo de seis unidades motoras que consumen combustible químico (ATP) y usan bucles en forma de dedo dentro de un poro estrecho para sujetar y tirar de la cola proteica.
Por qué unos pocos bucles de agarre importan tanto
Cada una de las seis unidades motoras tiene un par de bucles de poro que se introducen en el túnel central como ganchos y contactan la cadena proteica que pasa. Instantáneas estructurales anteriores mostraron estos ganchos dispuestos en una escalera en espiral alrededor del túnel, tomando turnos para agarrar y tirar. Pero no todos los ganchos parecían iguales. Para poner esto a prueba, los investigadores debilitaron un aminoácido clave en el bucle pore-1 de cada unidad motora en proteasomas de levadura y luego midieron qué tan bien la máquina consumía combustible, cambiaba de forma y degradaba proteínas modelo.
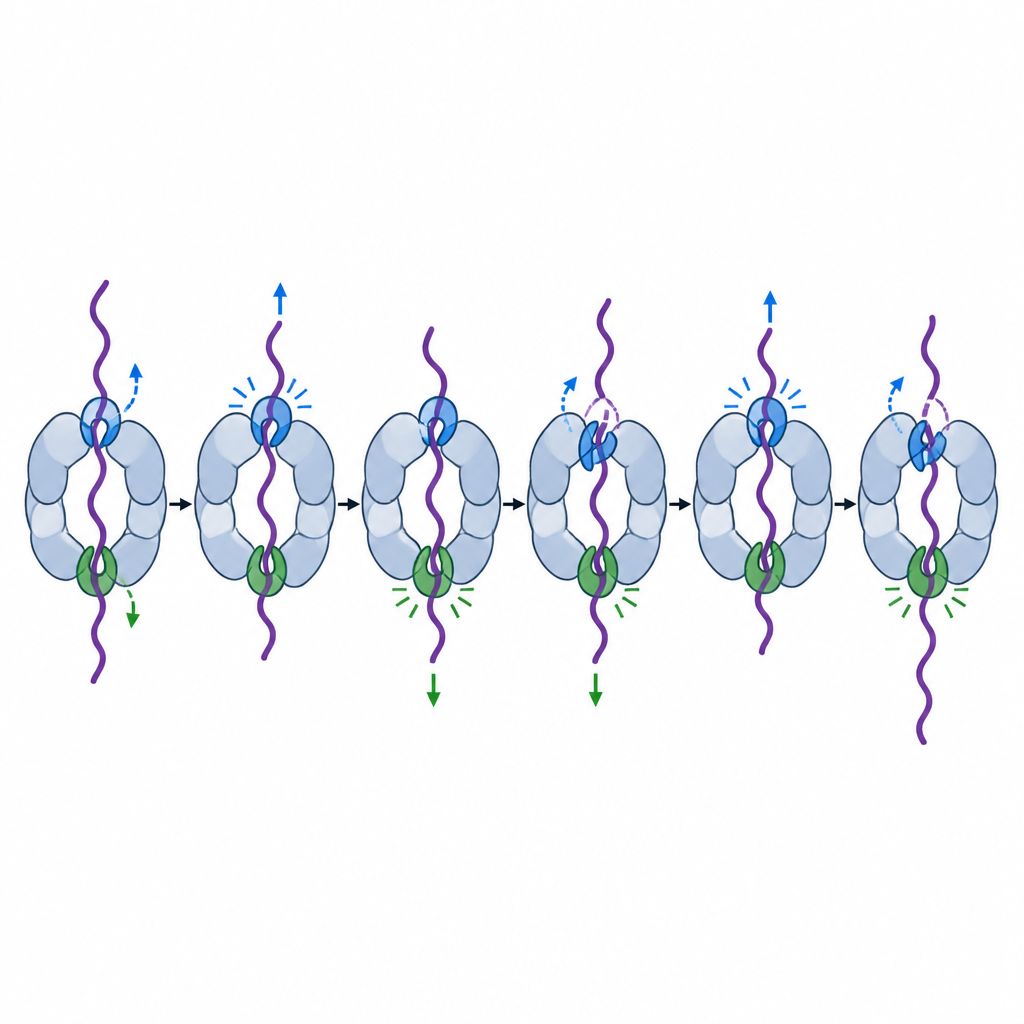
Dos ganchos especiales que guían la captura y el desenrollado
Combinando ensayos bioquímicos de conjunto, seguimiento fluorescente de molécula única y criomicroscopía electrónica de alta resolución, el equipo encontró que los bucles de poro en dos unidades motoras particulares, llamadas Rpt6 y Rpt4, desempeñan papeles especialmente importantes pero distintos. Cuando se debilitó el bucle Rpt6, el proteasoma consumía ATP incluso sin carga y pasaba más tiempo en una postura de “procesamiento” que bloquea parcialmente la entrada. Este mutante con frecuencia no lograba capturar firmemente las colas proteicas entrantes e incluso después de comenzar a trabajar sobre una proteína estable, seguía resbalando y tardando más o, en última instancia, permitía que el sustrato escapara. Las imágenes de crio-EM revelaron por qué: en el estado de reposo, el bucle Rpt6 está recogido en una forma helicoidal y se mantiene en su sitio mediante contactos inusuales con una subunidad vecina, aparentemente bloqueando la máquina en una configuración tranquila y lista para enganchar hasta que llega una proteína.
Mantener un agarre firme durante trabajos difíciles
El bucle Rpt4 tenía una especialidad diferente. Los proteasomas con un gancho Rpt4 debilitado aún podían reconocer y unirse a proteínas etiquetadas, pero cuando intentaban desenrollar un dominio proteico particularmente resistente, con frecuencia resbalaban y lo liberaban en lugar de tirarlo hasta el final. Trazas de molécula única mostraron intentos repetidos de desenrollar la misma proteína, interrumpidos por breves retornos al estado relajado, como si la máquina perdiera momentáneamente el agarre y tuviera que comenzar de nuevo. Comparaciones estructurales con vistas anteriores en escalera en espiral sugieren que Rpt4 a menudo ocupa una posición clave de “costura” justo antes de una carrera de fuerza, lo que lo convierte en el primer gancho que se cierra durante un empujón de tracción fuerte.
Un motor asimétrico afinado para la fiabilidad
En conjunto, los resultados presentan el motor del proteasoma como un motor asimétrico más que como un rotor perfectamente equilibrado de seis partes. Rpt6 ayuda a detectar cuando una proteína está en posición y desencadena el cambio de una postura de espera a una de trabajo, además de volver a enganchar la cadena tras deslizamientos ocasionales. Rpt4, a su vez, proporciona gran parte de la fuerza de tracción necesaria para desenrollar proteínas tercas sin soltarlas. Al asignar estas tareas distintas a diferentes ganchos alrededor del anillo, el proteasoma puede evitar desperdiciar energía y garantizar que, una vez que una proteína es seleccionada para la destrucción, por lo general se descomponga por completo en lugar de liberarse parcialmente.
Cita: López-Alfonzo, E., Saurabh, A., Zarafshan, S. et al. Substrate-interacting pore loops of two ATPase subunits determine the degradation efficiency of the 26S proteasome. Nat Commun 17, 4473 (2026). https://doi.org/10.1038/s41467-026-70426-y
Palabras clave: proteasoma, degradación de proteínas, motor ATPasa, sistema de ubiquitina, FRET de molécula única