Clear Sky Science · fr
Des boucles de pore en interaction avec le substrat de deux sous-unités ATPase déterminent l’efficacité de dégradation du protéasome 26S
Comment la machine de recyclage de la cellule agrippe les protéines endommagées
Chaque cellule doit en permanence éliminer les protéines endommagées ou devenues inutiles. Une immense machine moléculaire appelée le protéasome accomplit ce travail en aspirant les protéines, en les déchiquetant, puis en relâchant les fragments. Cette étude pose une question étonnamment précise aux conséquences importantes : comment deux minuscules éléments de préhension à l’intérieur du protéasome décident-ils si une protéine sera détruite efficacement ou pourra s’échapper ?
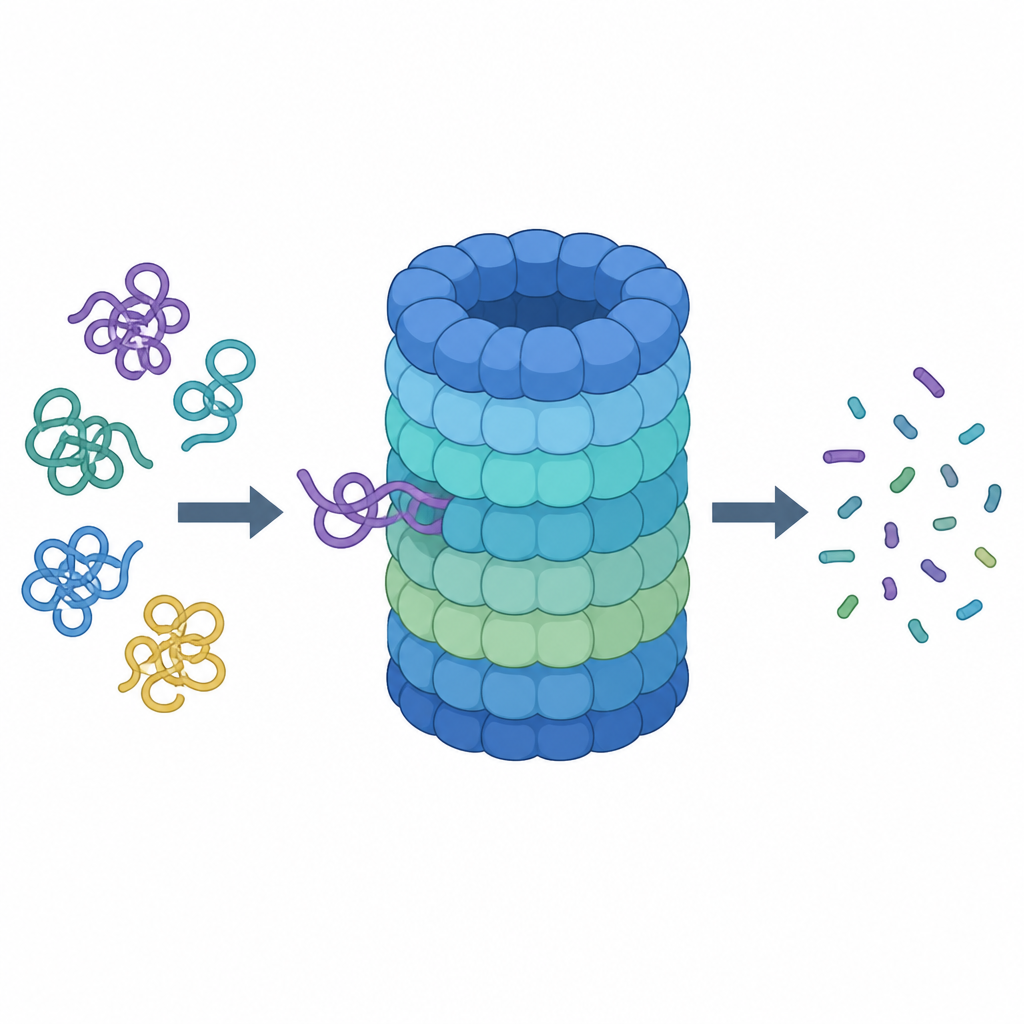
Un regard rapproché sur le broyeur protéique de la cellule
Le protéasome est une machine en forme de tonneau composée de nombreuses parties protéiques. Son tonneau central est la chambre de découpe, tandis qu’un chapeau au sommet reconnaît les protéines marquées pour destruction, enlève leurs « étiquettes » moléculaires et les introduit à l’intérieur. Pour être acceptée comme déchet, une protéine doit porter des chaînes d’une petite molécule de marquage appelée ubiquitine et exposer une queue flexible qui peut être agrippée et tirée. À l’entrée du capuchon se trouve un anneau de six unités motrices qui brûlent du carburant chimique (ATP) et utilisent des boucles en forme de doigts dans un pore étroit pour saisir et tirer sur la queue protéique.
Pourquoi quelques boucles de préhension ont autant d’importance
Chacune des six unités motrices possède une paire de boucles de pore qui s’étendent dans le tunnel central comme des crochets et contactent la chaîne protéique qui passe. Des instantanés structuraux antérieurs montraient ces crochets disposés en un escalier en spirale autour du tunnel, se relayant pour saisir et tirer. Mais tous les crochets ne semblaient pas équivalents. Pour tester cela, les chercheurs ont affaibli un acide aminé clé dans la boucle pore-1 de chaque unité motrice dans des protéasomes de levure, puis ont mesuré la capacité de la machine à brûler du carburant, à modifier sa conformation et à découper des protéines modèles.
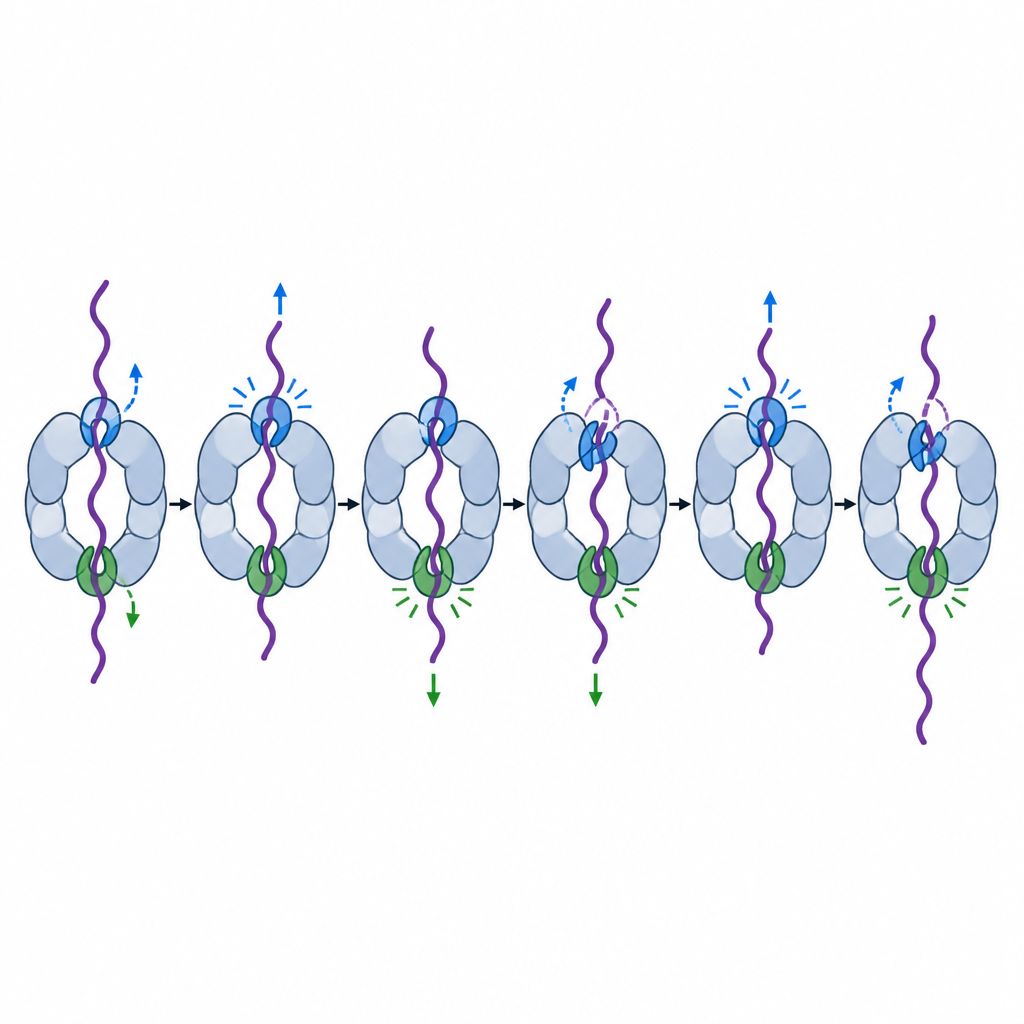
Deux crochets spéciaux qui guident la capture et le déroulement
En combinant des tests biochimiques en vrac, le suivi fluorescent moléculaire unique et la cryo-microscopie électronique à haute résolution, l’équipe a découvert que les boucles de pore de deux unités motrices particulières, appelées Rpt6 et Rpt4, jouent des rôles particulièrement importants mais distincts. Lorsque la boucle Rpt6 était affaiblie, le protéasome hydrolysait de l’ATP même en l’absence de cargaison et restait plus longtemps dans une posture de « traitement » qui bloque partiellement l’entrée. Ce mutant échouait souvent à capturer fermement les queues protéiques entrantes, et même après avoir commencé à travailler sur une protéine stable, continuait à glisser, prenant du retard ou finissant par laisser échapper le substrat. Des images en cryo-EM ont révélé la raison : à l’état repos, la boucle Rpt6 est repliée en une hélice et maintenue en place par des contacts inhabituels avec une sous-unité voisine, verrouillant apparemment la machine dans une configuration silencieuse prête à s’engager jusqu’à l’arrivée d’une protéine.
Maintenir une prise ferme lors des tâches difficiles
La boucle Rpt4 avait une spécialité différente. Les protéasomes portant un crochet Rpt4 affaibli pouvaient encore reconnaître et lier des protéines marquées, mais lorsqu’ils essayaient de dérouler un domaine protéique particulièrement robuste, ils glissaient fréquemment et le relâchaient au lieu de le tirer entièrement. Les traces monocellulaires ont montré des tentatives répétées pour dérouler la même protéine, interrompues par de brèves retours à l’état relâché, comme si la machine perdait momentanément sa prise et devait recommencer. Des comparaisons structurelles avec des vues antérieures en escalier en spirale suggèrent que Rpt4 occupe souvent une position clef de « couture » juste avant un coup de force, en faisant le premier crochet à se verrouiller lors d’une forte étape de traction.
Un moteur asymétrique réglé pour la fiabilité
Dans l’ensemble, les résultats dépeignent le moteur du protéasome comme un moteur asymétrique plutôt que comme un rotor parfaitement uniforme à six éléments. Rpt6 aide à détecter quand une protéine est en place et déclenche le passage d’une posture d’attente à une posture de travail, tout en ré-engageant la chaîne après des glissements occasionnels. Rpt4, à son tour, fournit une grande partie de la puissance de traction nécessaire pour dérouler des protéines récalcitrantes sans les lâcher. En confiant ces tâches distinctes à différents crochets autour de l’anneau, le protéasome peut à la fois éviter le gaspillage d’énergie et s’assurer que, une fois qu’une protéine est choisie pour destruction, elle est généralement entièrement dégradée plutôt que partiellement relâchée.
Citation: López-Alfonzo, E., Saurabh, A., Zarafshan, S. et al. Substrate-interacting pore loops of two ATPase subunits determine the degradation efficiency of the 26S proteasome. Nat Commun 17, 4473 (2026). https://doi.org/10.1038/s41467-026-70426-y
Mots-clés: protéasome, dégradation des protéines, moteur ATPase, système ubiquitine, FRET monoclonal