Clear Sky Science · it
I pori interagenti con il substrato di due subunità ATPasi determinano l’efficienza di degradazione del proteasoma 26S
Come la macchina di riciclo della cellula afferra le proteine danneggiate
Ogni cellula deve continuamente eliminare proteine danneggiate o non più necessarie. Una gigantesca macchina molecolare chiamata proteasoma svolge questo compito trascinando le proteine all’interno, sminuzzandole e rilasciando i frammenti. Questo studio affronta una domanda sorprendentemente specifica ma dalle grandi implicazioni: come fanno soltanto due piccolissime parti di presa all’interno del proteasoma a decidere se una proteina verrà distrutta in modo efficiente o potrà sfuggire?
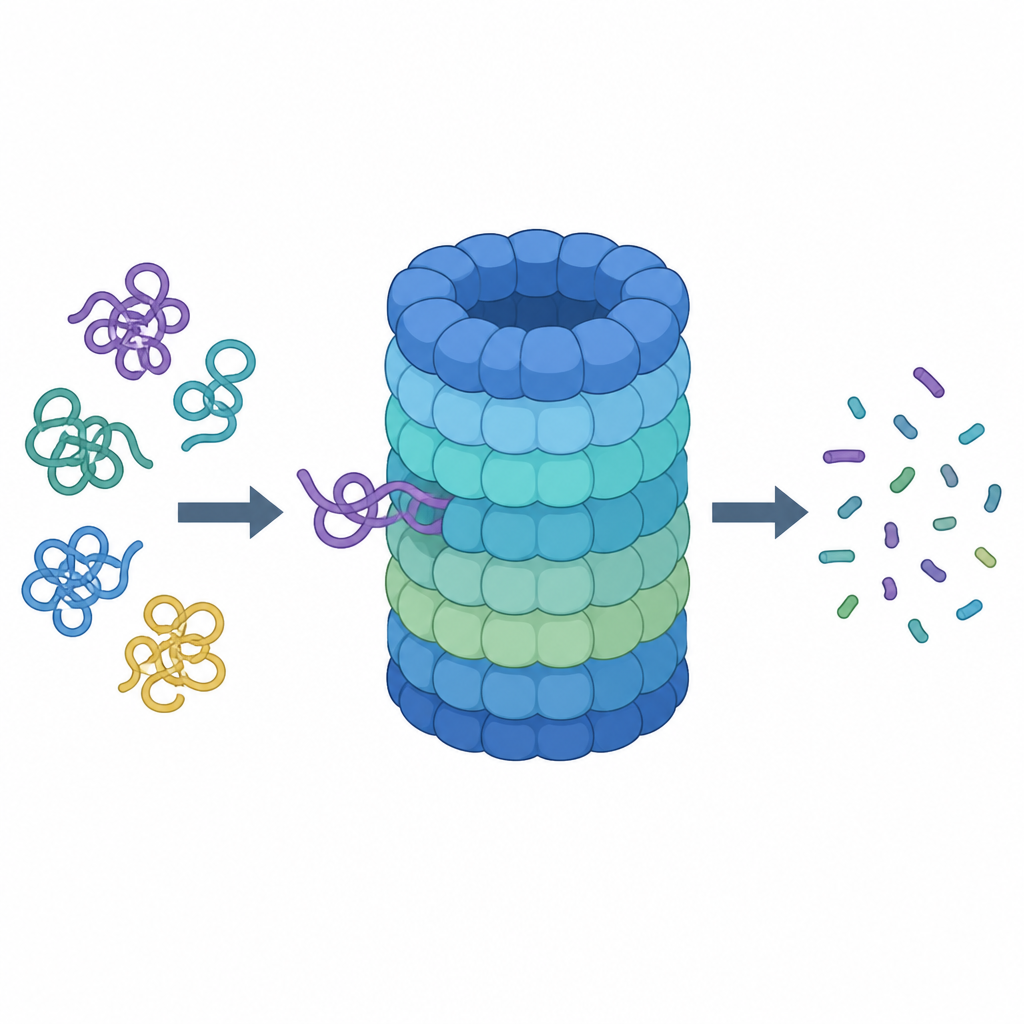
Uno sguardo ravvicinato al trituratore proteico della cellula
Il proteasoma è una macchina a forma di barile composta da molte subunità proteiche. Il suo barile centrale è la camera di taglio, mentre un cappuccio superiore riconosce le proteine marcate per la distruzione, rimuove i loro “tag” molecolari e le inserisce all’interno. Per essere accettata come rifiuto, una proteina deve portare catene di una piccola molecola di segnalazione chiamata ubiquitina ed esporre una coda floscia che può essere afferrata e tirata. All’ingresso del cappuccio si trova un anello di sei unità motorie che consumano carburante chimico (ATP) e usano anelli simili a dita all’interno di un poro stretto per impigliare e tirare la coda della proteina.
Perché pochi anelli di presa contano così tanto
Ciascuna delle sei unità motorie possiede una coppia di anelli di poro che si estendono nel tunnel centrale come uncini e vengono a contatto con la catena proteica che passa. Scatti strutturali precedenti mostravano questi uncini disposti a spirale intorno al tunnel, che a turno la afferravano e la tiravano. Ma non tutti gli uncini sembravano equivalenti. Per testarlo, i ricercatori indebolirono un amminoacido chiave nell’anello pore-1 di ciascuna unità motoria nei proteasomi di lievito, quindi misurarono quanto bene la macchina consumava carburante, cambiava forma e digeriva proteine modello.
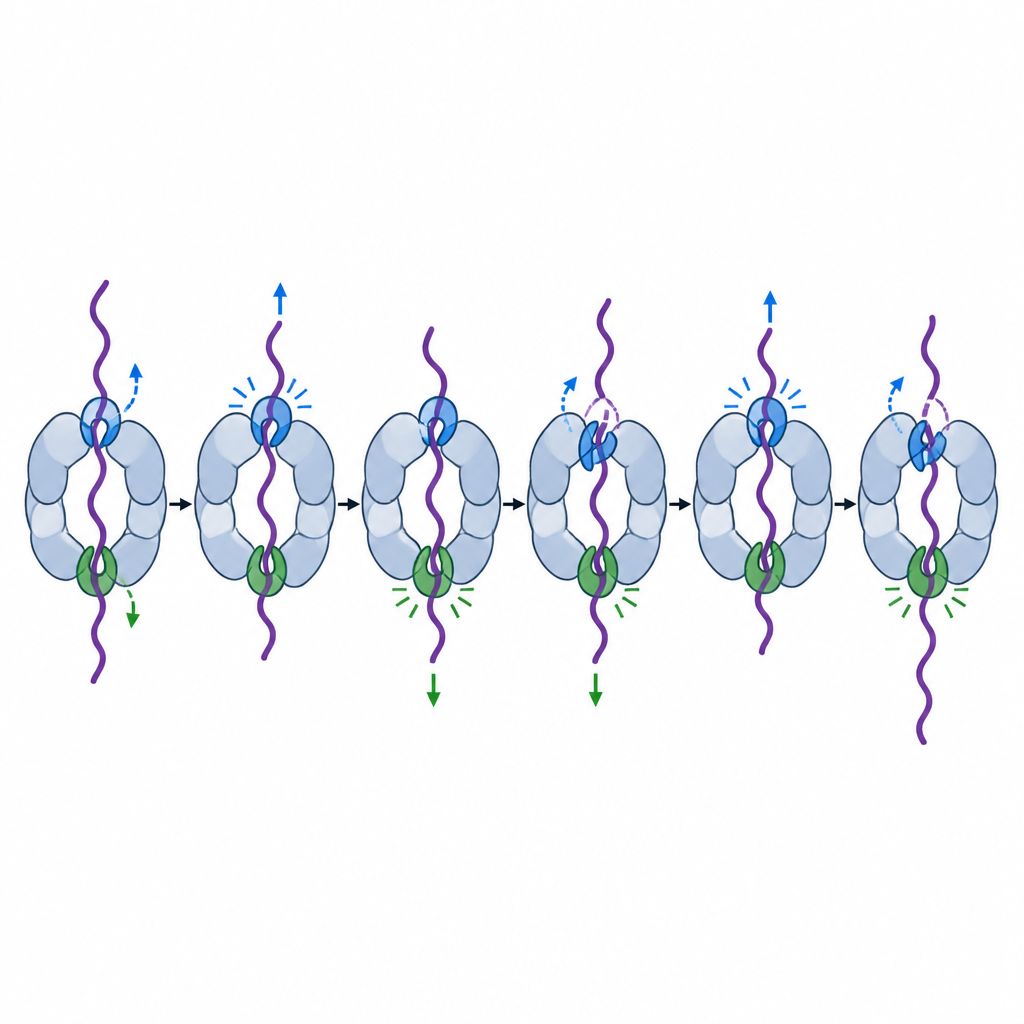
Due uncini speciali che guidano la cattura e il disordinamento
Combinando test biochimici su popolazioni, tracciamento di fluorescenza a molecola singola e criomicroscopia elettronica ad alta risoluzione, il gruppo ha scoperto che gli anelli di poro in due particolari unità motorie, chiamate Rpt6 e Rpt4, svolgono ruoli particolarmente importanti ma diversi. Quando l’anello Rpt6 veniva indebolito, il proteasoma consumava ATP anche in assenza di carico e trascorreva più tempo in una postura “di elaborazione” che ostruisce parzialmente l’ingresso. Questo mutante spesso non riusciva a catturare saldamente le code proteiche in entrata e, anche dopo aver iniziato a lavorare su una proteina stabile, continuava a slittare impiegando più tempo o permettendo infine al substrato di sfuggire. Immagini da cryo-EM hanno rivelato il perché: nello stato di riposo, l’anello Rpt6 è ripiegato in una forma elicoidale e mantenuto in posizione da contatti insoliti con una subunità vicina, apparentemente bloccando la macchina in una configurazione silenziosa e pronta all’ingaggio finché non arriva una proteina.
Mantenere una presa salda durante i compiti più difficili
L’anello Rpt4 aveva una specialità diversa. I proteasomi con un uncino Rpt4 indebolito potevano comunque riconoscere e legare proteine taggate, ma quando cercavano di disfarsi di un dominio proteico particolarmente robusto, spesso slittavano e lo rilasciavano invece di tirarlo completamente attraverso. I tracciati a molecola singola mostravano tentativi ripetuti di disfare la stessa proteina, interrotti da brevi ritorni allo stato rilassato, come se la macchina perdesse momentaneamente la presa e dovesse ricominciare. Confronti strutturali con viste a scala a chiocciola precedenti suggeriscono che Rpt4 occupa spesso una posizione chiave di “giunzione” subito prima di una corsa di potenza, rendendolo il primo uncino a stringersi durante una forte fase di trazione.
Un motore asimmetrico progettato per l’affidabilità
Nel complesso, i risultati dipingono il motore del proteasoma come un motore asimmetrico piuttosto che un rotore perfettamente uniforme in sei parti. Rpt6 aiuta a percepire quando una proteina è al suo posto e innesca il passaggio da una postura di attesa a una di lavoro, riallacciando anche la catena dopo scivolamenti occasionali. Rpt4, a sua volta, fornisce gran parte della forza di trazione necessaria a disfarsi di proteine ostinate senza lasciarle cadere. Assegnando questi compiti distinti a diversi uncini intorno all’anello, il proteasoma può sia evitare sprechi di energia sia garantire che, una volta scelta per la distruzione, una proteina venga generalmente scomposta completamente piuttosto che rilasciata parzialmente.
Citazione: López-Alfonzo, E., Saurabh, A., Zarafshan, S. et al. Substrate-interacting pore loops of two ATPase subunits determine the degradation efficiency of the 26S proteasome. Nat Commun 17, 4473 (2026). https://doi.org/10.1038/s41467-026-70426-y
Parole chiave: proteasoma, degradazione proteica, motore ATPasi, sistema dell’ubiquitina, FRET a molecola singola