Clear Sky Science · pl
Strukturalne podstawy promiskuityzmu niezwykłej ludzkiej hydroksylazy asparaginianu/asparaginy zależnej od Fe(II) i 2‑okso‑glutaranu
Dlaczego drobne zmiany w białkach mają znaczenie
W naszych komórkach funkcjonuje niezliczona liczba drobnych chemicznych modyfikacji białek, które utrzymują życie w prawidłowym stanie. Jedną z takich modyfikacji jest dodanie pojedynczego atomu tlenu do określonych reszt w białkach. W tej pracy skupiono się na ludzkim enzymie AspH, który przeprowadza tę subtelną przemianę i jest powiązany z nowotworami. Odkrywając sposób działania AspH na poziomie atomowym, badacze pokazują, jak może on działać na różne cele białkowe i sugerują nowe podejścia do projektowania leków, które selektywnie go wyłączają.
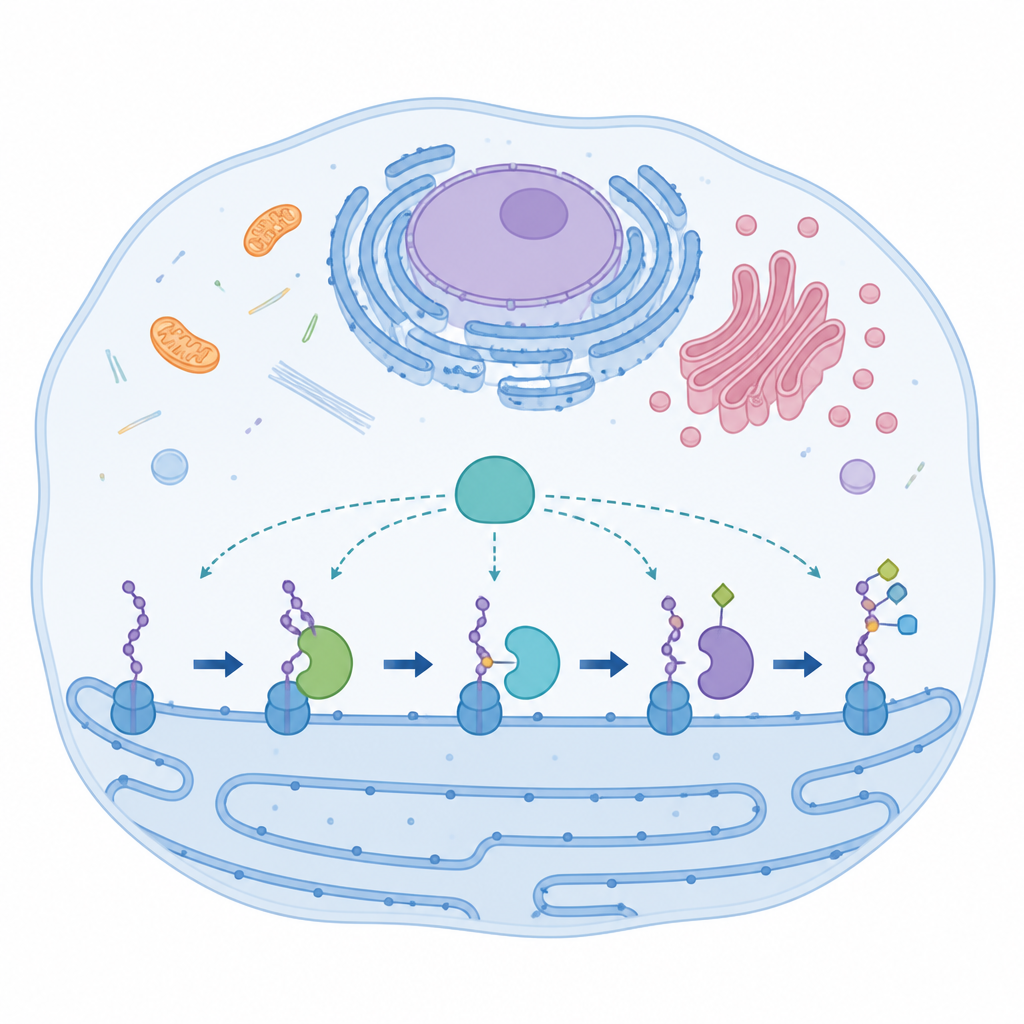
Specjalistyczny enzym z nietypowymi elementami
AspH działa w obrębie struktury komórkowej zwanej siateczką śródplazmatyczną, gdzie modyfikuje krótkie fragmenty białek znane jako domeny podobne do czynnika wzrostu naskórka (EGF‑like). Domeny te kontrolują procesy takie jak krzepnięcie krwi i sygnalizacja międzykomórkowa. Większość pokrewnych enzymów wykorzystuje standardowe trójszczękowe ułożenie ligandów metalu do utrzymania atomu żelaza napędzającego reakcję. AspH łamie tę zasadę: używa tylko dwóch bocznych łańcuchów histydyny z białka oraz ściśle związanej cząsteczki wody do uchwycenia żelaza. Pomimo tego nietypowego układu, AspH może działać zarówno na reszty asparaginianu, jak i asparaginy w swoich celach białkowych, co wskazuje na wbudowaną elastyczność, którą badacze określają jako promiskuityzm.
Obserwowanie reakcji w kryształach
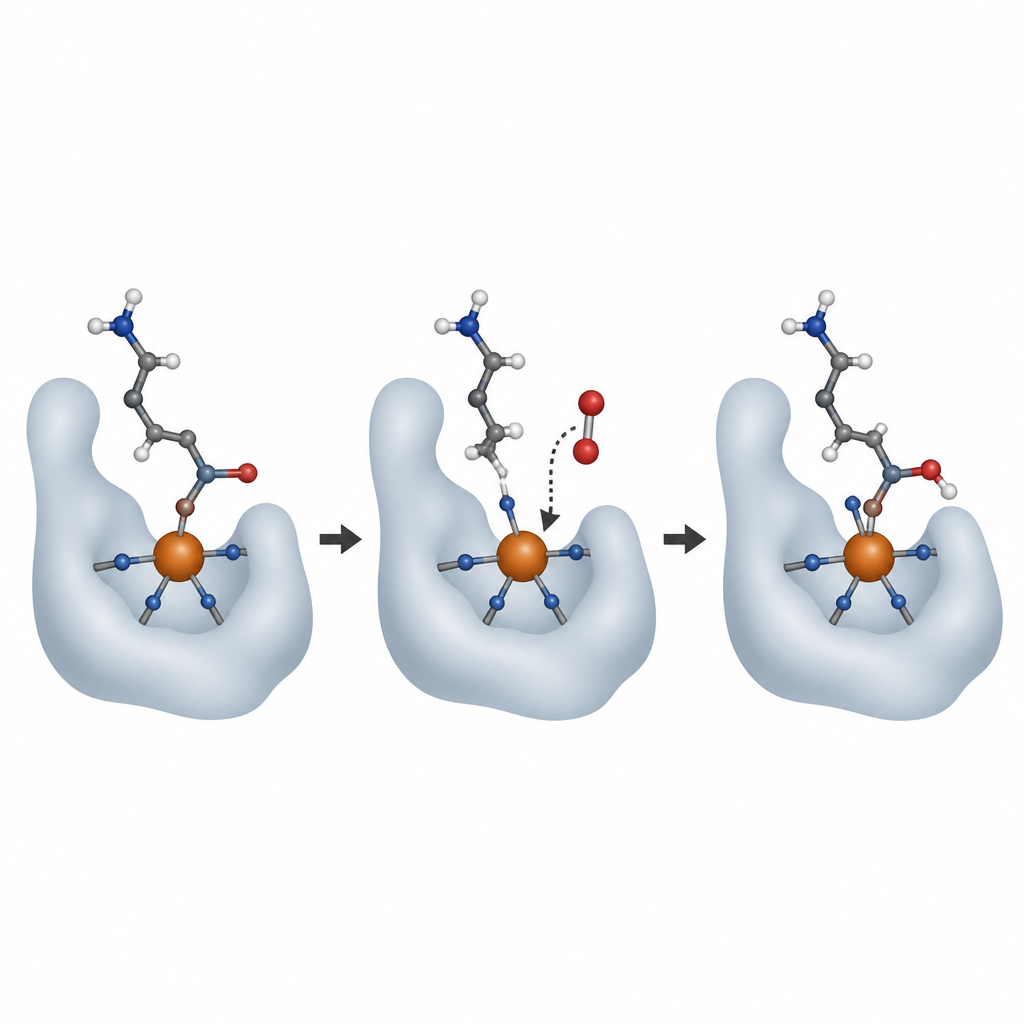
Aby zobaczyć AspH w akcji, zespół wyhodował kryształy aktywnego enzymu związane z koenzymem żelaza, pomocniczą cząsteczką 2‑okso‑glutaranu oraz krótkimi substratami białkowymi. Przy użyciu wysokoenergetycznych wiązek rentgenowskich w potężnych źródłach światła, w tym rentgenowskich laserów generujących wolne elektrony, uchwycono migawki enzymu przed i po reakcji z tlenem. W temperaturze pokojowej, nawet w sztywnej sieci krystalicznej, AspH przeprowadził pojedynczy cykl reakcji, dodając atom tlenu do substratu i przekształcając 2‑okso‑glutaran w bursztynian. Nowa grupa hydroksylowa produktu obróciła się, by związać żelazo, zajmując miejsce cząsteczki wody, która w stanie wyjściowym znajdowała się po przeciwnej stronie jednej z histydyn.
Jak tlen znajduje swoje miejsce
Naukowcy pytali następnie, gdzie przyłącza się nadchodzący tlen i jak zmienia się żelazo podczas reakcji. Użyli tlenku azotu, bliskiego odpowiednika O2, który można śledzić za pomocą spektroskopii rezonansu paramagnetycznego elektronów (EPR), jako substytutu dla O2. Zarówno w kryształach, jak i w roztworze tlenek azotu przyłączał się do żelaza w tym samym miejscu, które wcześniej zajmowała słabo związana cząsteczka wody. Dodatkowe pomiary emisji rentgenowskiej wykazały, że po pełnym obrocie żelazo wraca do pierwotnego stanu Fe(II), zgodnie z klasycznym cyklem, w którym żelazo przechodzi przejściowo przez wyższe stany utlenienia, napędzając reakcję, a następnie zostaje zresetowane do kolejnego cyklu. Starannie zaprojektowane eksperymenty z ciężkim tlenem (18O2) potwierdziły, że atom tlenu dodany do białka pochodzi bezpośrednio z tlenu cząsteczkowego, a nie z wody, mimo że cząsteczka wody jest zawsze związana z metalem.
Dostrajanie i granice elastyczności enzymu
Subtelne zmiany w otaczającej sieci wiązań wodorowych umożliwiają AspH obsługę zarówno asparaginianu, jak i asparaginy, choć nie z jednakową wydajnością. Elastyczny boczny łańcuch glutaminy (Q627) przesuwa się, by wchodzić w różne interakcje z każdym typem substratu i pobliskimi cząsteczkami wody, nieznacznie zmieniając efektywność reakcji. Zespół przetestował także pseudohalogen, izotiocyjanian, który może zachowywać się jak aniony halogenkowe wykorzystywane przez inne pokrewne enzymy do wprowadzania atomów chloru lub bromu zamiast tlenu. Izotiocyjanian wiązał się z żelazem AspH, ale w miejscu, które nie sprzyja chemii halogenacji. To niewłaściwe ulokowanie prawdopodobnie wyjaśnia, dlaczego AspH nie przeprowadza halogenacji, mimo że jego miejsce żelazowe przypomina to znane z enzymów halogenaz.
Co to znaczy dla chorób i terapii
AspH jest często nadmiernie produkowany i nieprawidłowo lokalizowany na powierzchni komórek w kilku typach nowotworów, gdzie jego aktywność wiąże się z większą inwazyjnością guzów i gorszym rokowaniem. Wyjaśniając, jak działa jego nietypowe, oparte na żelazie i wodzie miejsce metaliczne, badanie wskazuje nowe strategie projektowania leków. Zamiast jedynie naśladować pomocniczą cząsteczkę 2‑okso‑glutaran, przyszłe inhibitory mogłyby być zaprojektowane tak, by wypierać ściśle związaną wodę lub blokować miejsce wejścia tlenu, osiągając większą selektywność wobec AspH w porównaniu z innymi niezbędnymi ludzkimi enzymami. Zrozumienie tej delikatnie wyważonej chemii wspiera także hipotezę, że AspH może pomagać komórkom w wykrywaniu poziomu tlenu, oraz dostarcza strukturalnych podstaw do inżynierii pokrewnych enzymów dla nowych, bardziej ekologicznych reakcji chemicznych.
Cytowanie: de Munnik, M., Brasnett, A., Zhou, T. et al. Structural basis of the promiscuity of the unusual Fe(II) and 2-oxoglutarate dependent human aspartate/asparagine-β-hydroxylase. Nat Commun 17, 4267 (2026). https://doi.org/10.1038/s41467-026-69425-w
Słowa kluczowe: enzym AspH, hydroksylacja białek, tlenaza zależna od żelaza, wyczuwanie tlenu, biologia nowotworów