Clear Sky Science · es
Base estructural de la promiscuidad de la inusual aspartato/asparagina-β-hidroxilasa humana dependiente de Fe(II) y 2-oxoglutarato
Por qué importan los cambios diminutos en las proteínas
Nuestras células dependen de innumerables retoques químicos pequeños a las proteínas para mantener la vida en funcionamiento. Uno de esos ajustes consiste en añadir un solo átomo de oxígeno a ciertos componentes de las proteínas. Este estudio se centra en una enzima humana llamada AspH que realiza esta modificación sutil y que está relacionada con el cáncer. Al desvelar cómo funciona AspH a nivel atómico, los investigadores muestran cómo puede actuar sobre varios objetivos proteicos distintos y sugieren nuevas vías para diseñar fármacos que la inhiban selectivamente.
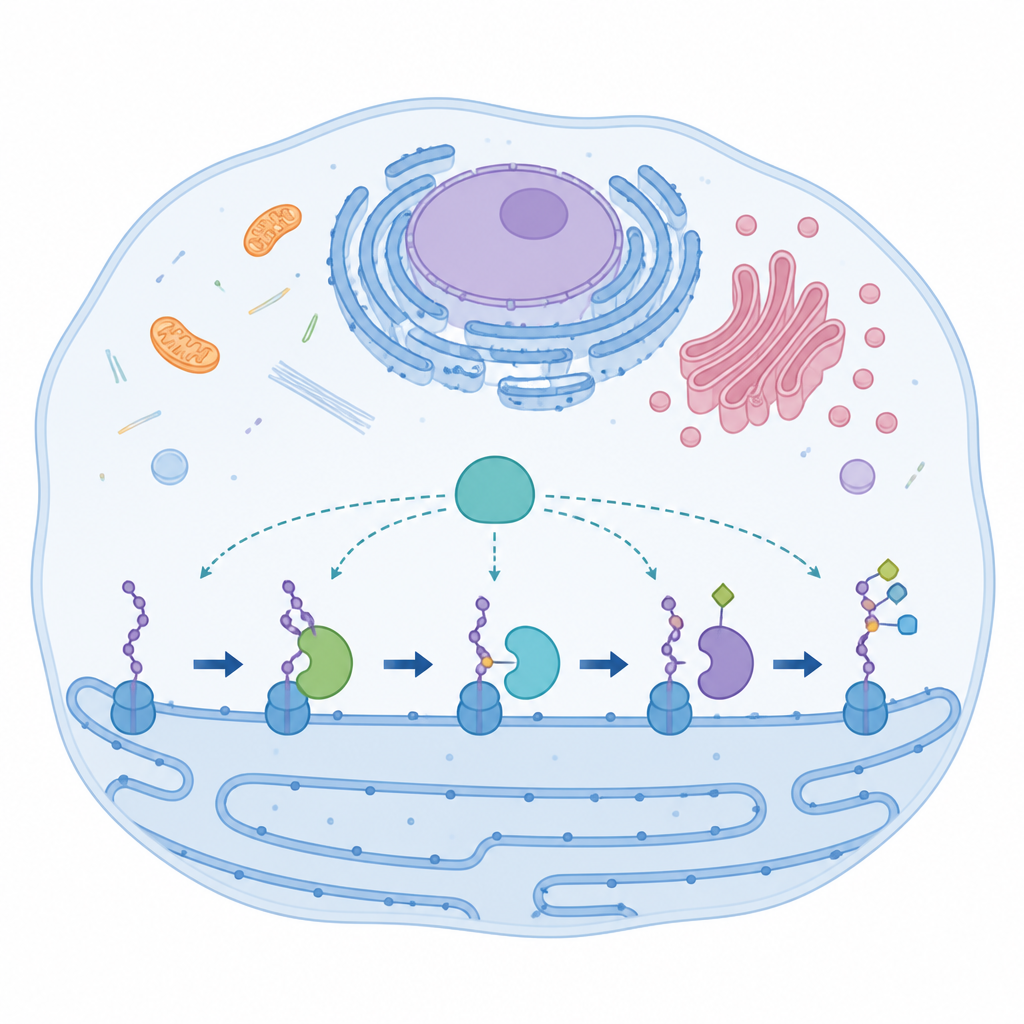
Una enzima especialista con partes inusuales
AspH actúa dentro de un compartimento celular llamado retículo endoplásmico, donde modifica segmentos cortos de proteínas conocidos como dominios tipo factor de crecimiento epidérmico. Estos dominios ayudan a controlar procesos como la coagulación sanguínea y la señalización celular. La mayoría de enzimas relacionadas utilizan una disposición habitual de tres ligandos para unir un átomo de hierro que impulsa la química. AspH rompe esta norma: emplea solo dos cadenas laterales de histidina de la proteína más una molécula de agua fuertemente ligada para sujetar el hierro. A pesar de esta configuración inusual, AspH puede actuar sobre residuos de aspartato y asparagina en sus dianas proteicas, lo que sugiere una flexibilidad intrínseca que los investigadores denominan promiscuidad.
Observando la química dentro de cristales
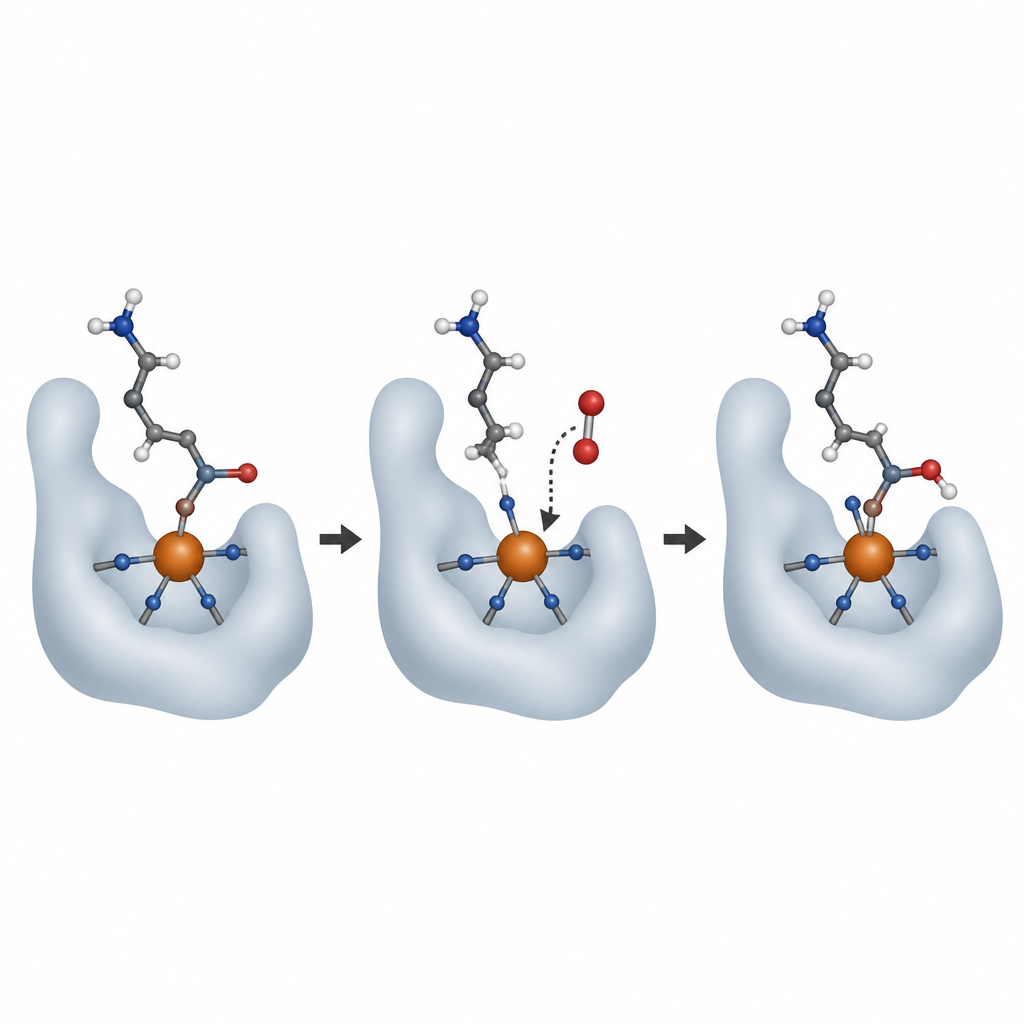
Para ver a AspH en acción, el equipo cristalizó la enzima activa unida a su cofactor de hierro, la molécula ayudante 2-oxoglutarato y sustratos proteicos cortos. Empleando haces de rayos X de alta intensidad en fuentes luminosas potentes, incluidos láseres de electrones libres de rayos X, capturaron instantáneas de la enzima antes y después de reaccionar con oxígeno. A temperatura ambiente e incluso dentro de la rígida red cristalina, AspH realizó una ronda de química, añadiendo un átomo de oxígeno al sustrato y convirtiendo el 2-oxoglutarato en succinato. El nuevo grupo hidroxilo del producto se giró para unirse al hierro, sustituyendo a una molécula de agua que en el estado inicial estaba opuesta a una de las histidinas.
Cómo encuentra su sitio el oxígeno
Los científicos se preguntaron entonces dónde se une el oxígeno entrante y cómo cambia el hierro durante la reacción. Utilizaron monóxido de nitrógeno, un análogo cercano del oxígeno que puede seguirse por espectroscopía de resonancia paramagnética electrónica, para representar al O2. Tanto en cristales como en solución, el monóxido de nitrógeno se unió al hierro en la misma posición ocupada previamente por esa agua débilmente ligada. Mediciones adicionales de emisión de rayos X mostraron que, tras una rotación completa, el hierro volvió a su estado original Fe(II), coherente con el ciclo clásico en el que el hierro pasa brevemente por estados de mayor energía para impulsar la reacción y luego se restablece para otra ronda. Experimentos diseñados con gas oxígeno pesado (18O2) confirmaron que el átomo de oxígeno añadido a la proteína procede directamente del oxígeno molecular, no del agua, a pesar de que una molécula de agua está siempre ligada al metal.
Ajustes finos y límites de la flexibilidad enzimática
Cambios sutiles en la red de enlaces de hidrógeno circundante permiten que AspH acomode tanto aspartato como asparagina, aunque no con la misma eficiencia. Una cadena lateral de glutamina flexible (Q627) cambia de posición para interactuar de forma distinta con cada tipo de sustrato y con las aguas próximas, alterando levemente la eficiencia de la reacción. El equipo también probó un pseudohaluro, isotiocianato, que puede comportarse como iones haluro que otras enzimas relacionadas usan para introducir átomos de cloro o bromo en lugar de oxígeno. El isotiocianato sí se unió al hierro de AspH, pero en un sitio que no favorece la química de halogenación. Esta mala colocación probablemente explica por qué AspH no realiza halogenación, aunque su sitio de hierro se asemeje al de enzimas halogenantes conocidas.
Qué significa esto para la enfermedad y la terapia
AspH se sobreexpresa con frecuencia y está mal localizada en la superficie celular en varios cánceres, donde su actividad se asocia con mayor invasividad tumoral y peores resultados. Al clarificar cómo funciona su inusual sitio metálico basado en hierro y agua, este estudio apunta a nuevas estrategias para el diseño de fármacos. En lugar de limitarse a imitar la molécula ayudante 2-oxoglutarato, futuros inhibidores podrían diseñarse para desplazar la molécula de agua fuertemente ligada o bloquear la entrada de oxígeno, logrando mayor selectividad por AspH frente a otras enzimas humanas esenciales. Comprender esta química finamente equilibrada también respalda la idea de que AspH podría ayudar a las células a detectar los niveles de oxígeno, y proporciona un marco estructural para ingeniería de enzimas relacionadas para nuevos tipos de reacciones químicas más sostenibles.
Cita: de Munnik, M., Brasnett, A., Zhou, T. et al. Structural basis of the promiscuity of the unusual Fe(II) and 2-oxoglutarate dependent human aspartate/asparagine-β-hydroxylase. Nat Commun 17, 4267 (2026). https://doi.org/10.1038/s41467-026-69425-w
Palabras clave: enzima AspH, hidroxilación de proteínas, oxigenasa dependiente de hierro, detección de oxígeno, biología del cáncer