Clear Sky Science · fr
Base structurale de la promiscuïté de l’étonnante aspartate/asparagine-β-hydroxylase humaine dépendante de Fe(II) et de 2‑oxoglutarate
Pourquoi de minuscules modifications protéiques comptent
Nos cellules dépendent d’innombrables ajustements chimiques mineurs des protéines pour maintenir le fonctionnement biologique. L’un de ces ajustements consiste à ajouter un atome d’oxygène à certaines unités constitutives des protéines. Cette étude se concentre sur une enzyme humaine nommée AspH qui réalise cette modification subtile et qui est liée au cancer. En dévoilant le fonctionnement atomique d’AspH, les chercheurs montrent comment elle peut agir sur plusieurs cibles protéiques et suggèrent de nouvelles voies pour concevoir des médicaments qui l’inhiberaient de façon sélective.
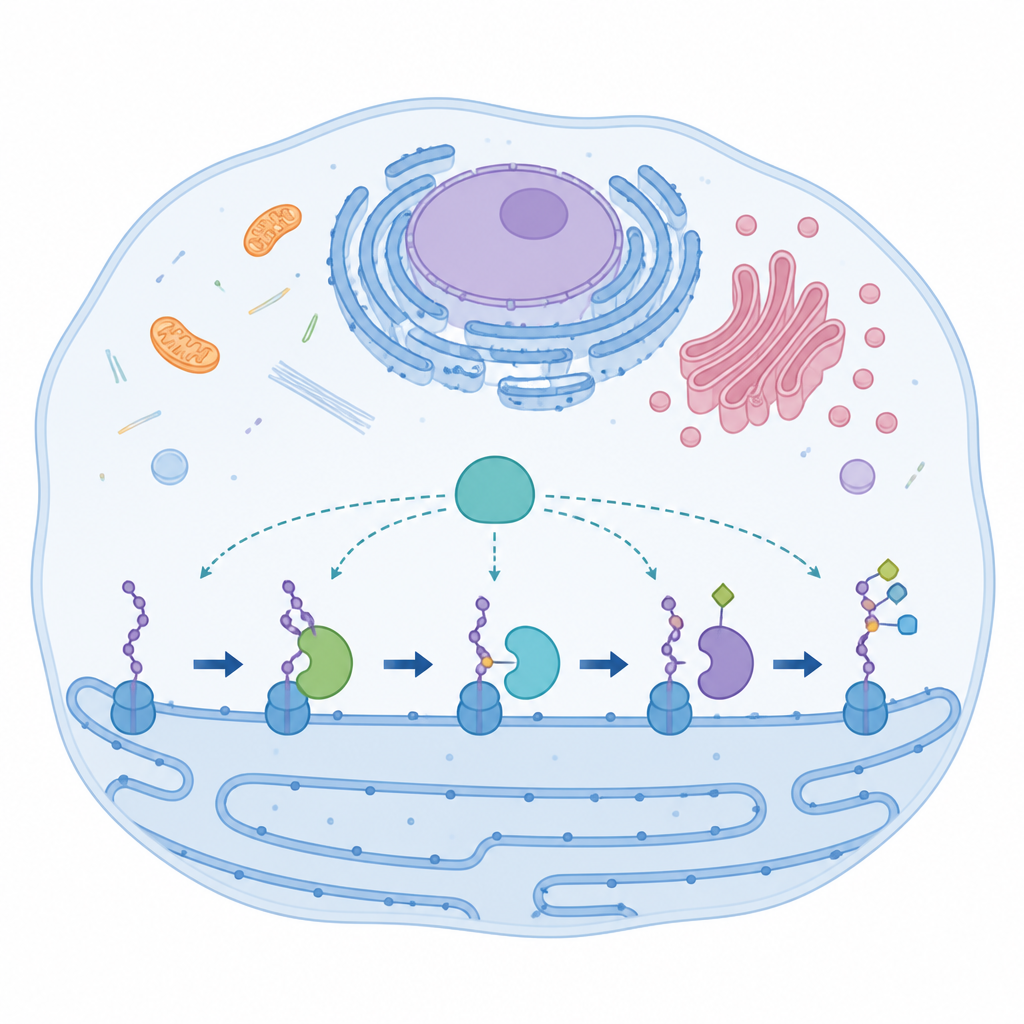
Une enzyme spécialisée aux éléments atypiques
AspH agit dans un compartiment cellulaire appelé réticulum endoplasmique, où elle modifie de courts segments protéiques connus sous le nom de domaines de type facteur de croissance épidermique. Ces domaines contribuent à réguler des processus tels que la coagulation et la signalisation cellulaire. La plupart des enzymes apparentées utilisent une coordination métallique classique en trois points pour maintenir un atome de fer qui catalyse la réaction. AspH rompt cette règle : elle n’emploie que deux chaînes latérales d’histidine de la protéine plus une molécule d’eau fermement tenue pour lier le fer. Malgré cette configuration inhabituelle, AspH peut agir sur des résidus d’aspartate comme d’asparagine dans ses cibles protéiques, ce qui suggère une flexibilité intégrée que les chercheurs qualifient de promiscuïté.
Observer la chimie à l’intérieur des cristaux
Pour voir AspH en action, l’équipe a fait croître des cristaux de l’enzyme active liée à son cofacteur fer, au co-substrat 2‑oxoglutarate et à de courts substrats protéiques. À l’aide de faisceaux de rayons X à haute intensité dans des sources lumineuses puissantes, y compris des lasers à électrons libres, ils ont capturé des instantanés de l’enzyme avant et après réaction avec l’oxygène. À température ambiante et même au sein du réseau cristallin rigide, AspH a effectué un cycle unique de réaction, ajoutant un atome d’oxygène au substrat et convertissant le 2‑oxoglutarate en succinate. Le nouveau groupe hydroxyle du produit s’est réorienté pour se lier au fer, prenant la place d’une molécule d’eau qui se trouvait en vis‑à‑vis d’une des histidines à l’état initial.
Comment l’oxygène trouve sa place
Les scientifiques ont ensuite cherché où se fixe l’oxygène entrant et comment le fer évolue durant la réaction. Ils ont utilisé le monoxyde d’azote, un analogue proche de l’oxygène traçable par spectroscopie par résonance paramagnétique électronique, pour se substituer à O2. Dans les cristaux comme en solution, le monoxyde d’azote s’est attaché au fer à la même position précédemment occupée par cette eau faiblement liée. Des mesures supplémentaires d’émission X ont montré qu’après un cycle complet, le fer revenait à son état initial Fe(II), conforme au cycle classique où le fer traverse brièvement des états d’oxydation supérieurs pour conduire la réaction puis se réinitialise pour un nouveau tour. Des expériences soigneusement conçues avec du gaz oxygène lourd (18O2) ont confirmé que l’atome d’oxygène ajouté à la protéine provient directement de l’oxygène moléculaire, et non de l’eau, même si une molécule d’eau reste toujours liée au métal.
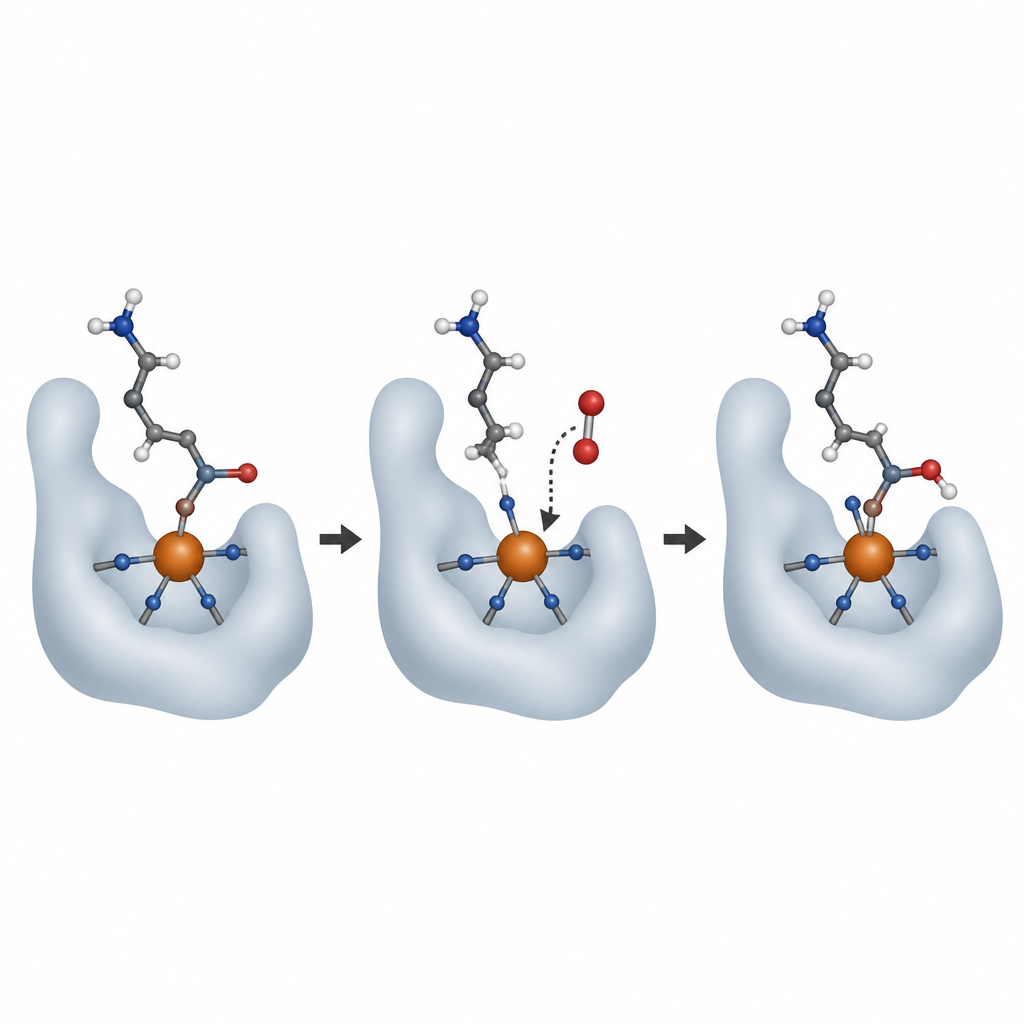
Ajustements fins et limites de la flexibilité enzymatique
Des changements subtils dans le réseau de liaisons hydrogène environnant permettent à AspH de prendre en charge soit l’aspartate soit l’asparagine, mais pas avec la même efficacité. Une chaîne latérale de glutamine flexible (Q627) change de position pour interagir différemment avec chaque type de substrat et avec les eaux voisines, modifiant légèrement l’efficacité de la réaction. L’équipe a aussi testé un pseudohalogénure, l’isothiocyanate, qui peut se comporter comme des ions halogénures que d’autres enzymes apparentées utilisent pour installer des atomes de chlore ou de brome au lieu d’oxygène. L’isothiocyanate s’est bien lié au fer d’AspH, mais en un site qui ne permet pas la chimie d’halogénation. Ce mauvais positionnement explique probablement pourquoi AspH n’effectue pas d’halogénation, bien que son site ferrique ressemble à celui d’enzymes halogénantes connues.
Implications pour la maladie et la thérapie
AspH est fréquemment surexprimée et mal localisée à la surface cellulaire dans plusieurs cancers, où son activité est associée à une plus grande invasivité tumorale et à un pronostic plus défavorable. En clarifiant le fonctionnement de son site métallique inhabituel basé sur le fer et l’eau, cette étude ouvre de nouvelles stratégies pour la conception de médicaments. Plutôt que de seulement imiter le co‑substrat 2‑oxoglutarate, de futurs inhibiteurs pourraient être conçus pour déplacer l’eau fermement tenue ou bloquer le site d’entrée de l’oxygène, obtenant ainsi une plus grande sélectivité pour AspH par rapport aux autres enzymes humaines essentielles. Comprendre cette chimie finement équilibrée soutient aussi l’idée qu’AspH pourrait aider les cellules à détecter les niveaux d’oxygène, et fournit un cadre structural pour l’ingénierie d’enzymes apparentées pour de nouveaux types de réactions chimiques « vertes ».
Citation: de Munnik, M., Brasnett, A., Zhou, T. et al. Structural basis of the promiscuity of the unusual Fe(II) and 2-oxoglutarate dependent human aspartate/asparagine-β-hydroxylase. Nat Commun 17, 4267 (2026). https://doi.org/10.1038/s41467-026-69425-w
Mots-clés: enzyme AspH, hydroxylation des protéines, oxygénase dépendante du fer, détection de l’oxygène, biologie du cancer