Clear Sky Science · nl
Structurele basis van de promiscuïteit van de ongebruikelijke Fe(II)- en 2‑oxoglutarine‑afhankelijke menselijke aspartaat/asparagine-β-hydroxylase
Waarom kleine eiwitwijzigingen ertoe doen
Onze cellen zijn afhankelijk van talloze kleine chemische aanpassingen aan eiwitten om het leven soepel te laten verlopen. Een van die aanpassingen is het toevoegen van een enkele zuurstofatoom aan bepaalde aminozuurgroepen binnen eiwitten. Deze studie richt zich op een menselijk enzym genaamd AspH dat deze subtiele modificatie uitvoert en een verband met kanker heeft. Door te onthullen hoe AspH op atomair niveau werkt, laten de onderzoekers zien hoe het op verschillende eiwitdoelen kan inwerken en suggereren ze nieuwe manieren om geneesmiddelen te ontwerpen die het selectief uitschakelen.
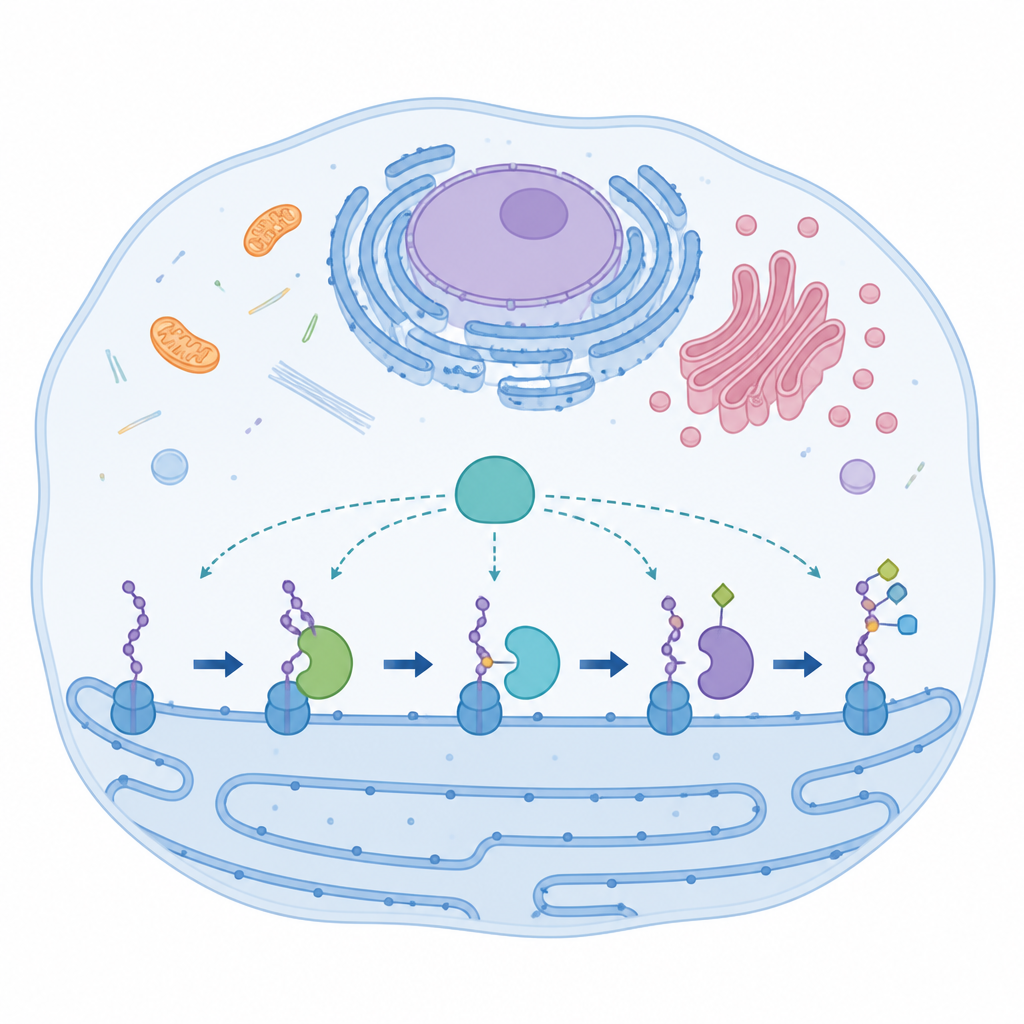
Een specialistisch enzym met ongebruikelijke onderdelen
AspH werkt in een celcompartiment dat het endoplasmatisch reticulum wordt genoemd, waar het korte eiwitsegmenten modificeert die bekendstaan als epidermal growth factor‑achtige domeinen. Deze domeinen helpen processen te regelen zoals bloedstolling en celsignalering. De meeste verwante enzymen gebruiken een standaard driedelige metaalbindingsorde om een ijzeratoom vast te houden dat de chemie aandrijft. AspH doorbreekt deze regel: het gebruikt slechts twee histidinezijgroepen van het eiwit plus een strak gebonden watermolecuul om het ijzer te grijpen. Ondanks deze ongebruikelijke opstelling kan AspH zowel aspartaat- als asparagine-residuen in zijn eiwitdoelen bewerken, wat wijst op een ingebouwde flexibiliteit die de onderzoekers promiscuïteit noemen.
De chemie in kristallen volgen
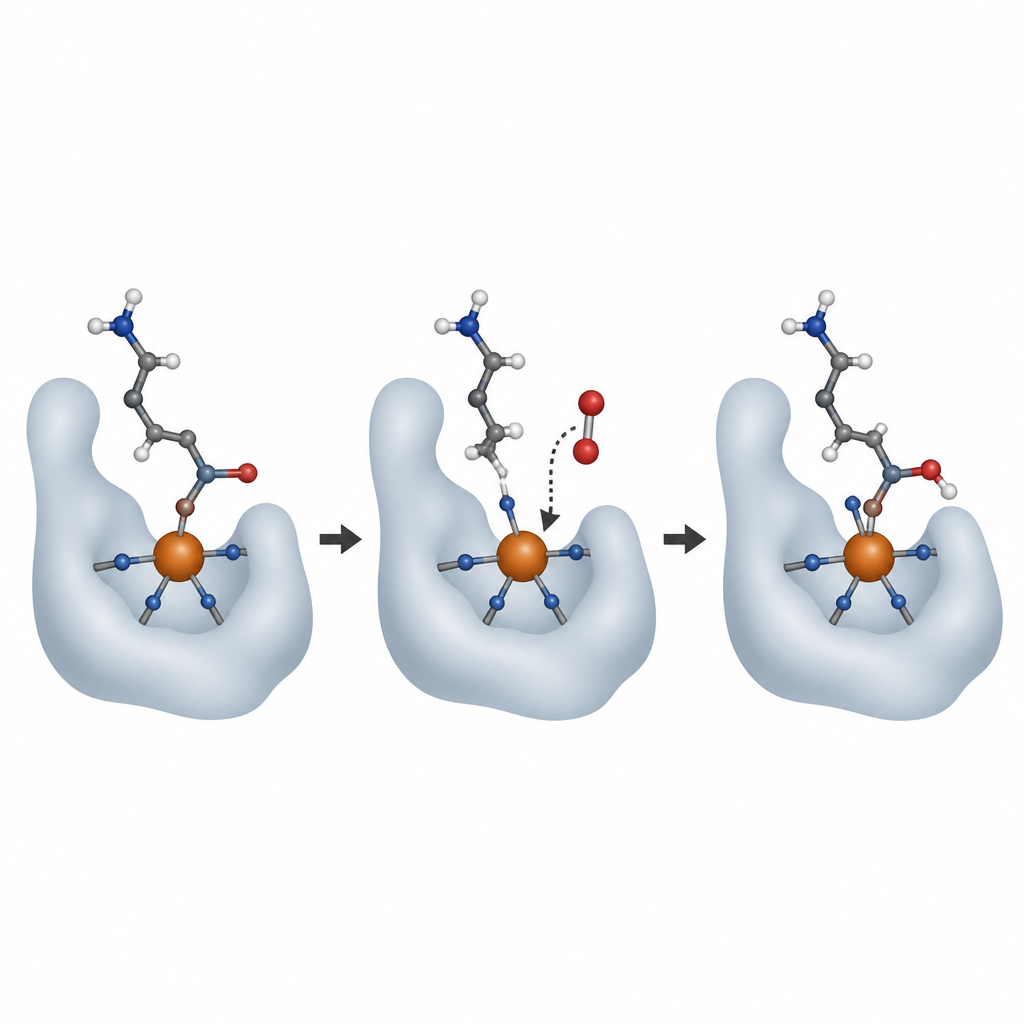
Om AspH in actie te zien, kweekte het team kristallen van het actieve enzym gebonden aan zijn ijzercofactor, het hulpstofmolecuul 2‑oxoglutarine, en korte eiwitsubstraten. Met behulp van hoogintensieve röntgenbundels bij krachtige lichtbronnen, waaronder röntgen‑vrije‑elektronlasers, legden ze momentopnames vast van het enzym voor en na reactie met zuurstof. Bij kamertemperatuur en zelfs binnen het starre kristalrooster voerde AspH één reactieronde uit, waarbij een zuurstofatoom aan het substraat werd toegevoegd en 2‑oxoglutarine werd omgezet in succinaat. De nieuwe hydroxylgroep van het product draaide om om het ijzer te binden en nam de plaats in van een watermolecuul dat tegenover een van de histidines in de begintoestand zat.
Hoe zuurstof zijn plaats vindt
Vervolgens vroegen de wetenschappers waar binnenkomende zuurstof bindt en hoe het ijzer tijdens de reactie verandert. Ze gebruikten stikstofoxide, een nauwe nabootser van zuurstof die kan worden gevolgd met elektronparamagnetische resonantiespectroscopie, als plaatsvervanger voor O2. Zowel in kristallen als in oplossing hechtte stikstofoxide zich aan het ijzer op dezelfde plaats die eerder werd ingenomen door dat zwak gebonden water. Aanvullende röntgenemissiemetingen toonden dat het ijzer na een volledige omzetting terugkeerde naar zijn oorspronkelijke Fe(II)-toestand, wat consistent is met de klassieke cyclus waarbij ijzer tijdelijk hogere energietoestanden doorloopt om de reactie aan te drijven en zich daarna reset voor een volgende ronde. Zorgvuldig ontworpen experimenten met zwaar zuurstofgas (18O2) bevestigden dat het zuurstofatoom dat aan het eiwit wordt toegevoegd rechtstreeks afkomstig is van molecuulzuurstof en niet van water, ondanks dat er altijd een watermolecuul aan het metaal gebonden is.
Fijnafstelling en grenzen van enzymflexibiliteit
Subtiele veranderingen in het omliggende waterstofbindingsnetwerk stellen AspH in staat zowel aspartaat als asparagine te verwerken, maar niet even efficiënt. Een flexibele glutaminezijgroep (Q627) verschuift van positie om verschillend te interageren met elk type substraat en nabijgelegen wateren, wat de reactieweerstand iets verandert. Het team testte ook een pseudohalide, isothiocyanate, dat zich als halide-ionen kan gedragen die andere verwante enzymen gebruiken om chloor‑ of broomatomen te installeren in plaats van zuurstof. Isothiocyanate bond wel aan het ijzer van AspH, maar op een plek die halogenatiechemie niet ondersteunt. Deze verkeerde plaatsing verklaart waarschijnlijk waarom AspH geen halogenatie uitvoert, ook al lijkt zijn ijzerplaats op die van bekende halogeniserende enzymen.
Wat dit betekent voor ziekte en therapie
AspH wordt in verschillende vormen van kanker vaak overgeproduceerd en misgelokaliseerd naar het celoppervlak, waar zijn activiteit wordt gekoppeld aan grotere tumorinvasiviteit en slechtere prognoses. Door te verduidelijken hoe zijn ongebruikelijke ijzer- en watergebaseerde metaalplaats werkt, wijst deze studie op nieuwe strategieën voor geneesmiddeldesign. In plaats van alleen het hulpstofmolecuul 2‑oxoglutarine na te bootsen, zouden toekomstige remmers zo ontworpen kunnen worden dat ze het strak gebonden water verdringen of de zuurstoftoegang blokkeren, waarmee een grotere selectiviteit voor AspH ten opzichte van andere essentiële menselijke enzymen kan worden bereikt. Inzicht in deze fijn afgewogen chemie ondersteunt ook het idee dat AspH cellen kan helpen zuurstofniveaus waar te nemen, en het biedt een structureel kader voor het ontwerpen van verwante enzymen voor nieuwe vormen van milieuvriendelijke chemische reacties.
Bronvermelding: de Munnik, M., Brasnett, A., Zhou, T. et al. Structural basis of the promiscuity of the unusual Fe(II) and 2-oxoglutarate dependent human aspartate/asparagine-β-hydroxylase. Nat Commun 17, 4267 (2026). https://doi.org/10.1038/s41467-026-69425-w
Trefwoorden: AspH-enzym, eiwit-hydroxylatie, ijzerafhankelijke oxygenase, zuurstofwaarneming, kankerbiologie