Clear Sky Science · zh
非萼输入门控萼式终板引发的MNTB输出时间
大脑如何追踪声音来源
确定声音来自何处,比如黑暗中一声折断的树枝,依赖于大脑发出极其迅速且时序精确的信号。本研究考察了位于听觉脑干的小型中继站,显示一组被忽视的侧向输入可以微妙地重新调整传出信号的时序,帮助大脑构建可靠的空间声源图谱。
关键听觉中继的两类输入
在梯状体内侧核这一深处脑干结构中,主细胞既接受称为霍尔德萼式终板的大型主输入,也在胞体和树突上接收许多较小的“非萼式”输入。萼式终板传递非常大且快速的信号,但在快速重复活动中其强度会下降。相比之下,较小的输入起初较弱,但在反复激活时会增强,并以更分散、非同步的方式持续释放神经递质。研究者在沙鼠和厄特鲁斯坎鼩鼱的脑片中记录到这些信号,发现这种分工在这些相距甚远的哺乳动物物种中是保守的。 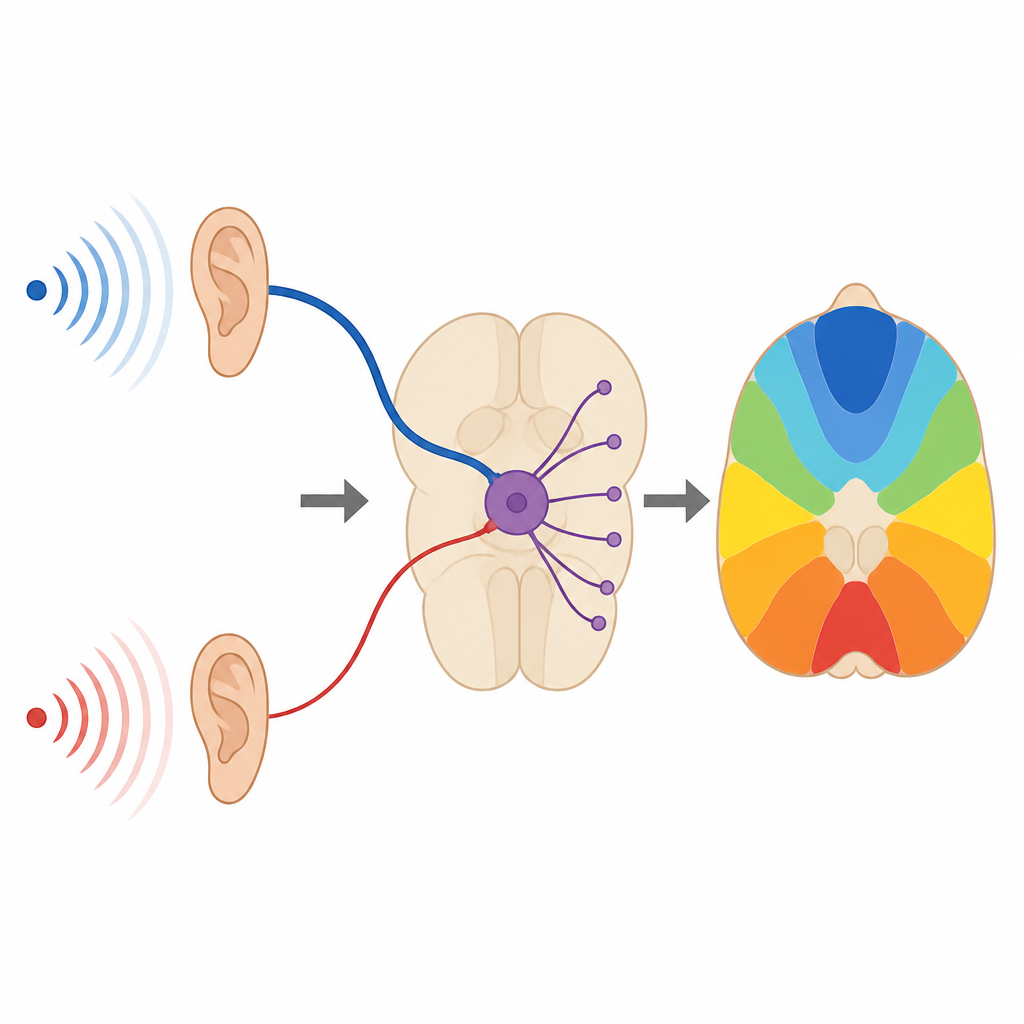
塑造持续活动的短期变化
团队关注短期可塑性,即在持续使用过程中突触的快速增强或减弱。他们表明,霍尔德萼式终板在高频刺激下出现强烈抑制,尤其在沙鼠中明显,每一次连续反应都会变小。相反,非萼式输入显示出显著的促进效应,随着刺激列车反应逐步增大,仅在极高频率下才轻微减弱。这些较小的输入还产生延迟事件,在刺激列车结束后持续数百毫秒,实际上提供的是一种稳定的去极化驱动,而不是精确定时的尖峰。提高灌流液中的钙浓度会使非萼式输入从促进转为抑制,并揭示它们的囊泡库以与萼式终板已知相似的时间常数补充。
模拟联合输入如何控制尖峰
为了测试这些不同输入如何协同工作,作者使用了动态钳,这是一种允许实验者向真实神经元注入计算机生成电导的技术。他们构建了模拟萼式和非萼式突触记录行为的模板,并有无模拟持续背景活动的预处理期。当将这些模板应用于MNTB神经元时,他们发现动作电位的发生概率、时机和精确度主要取决于到达胞体的总突触电导。单独的微小输入很少能驱动尖峰,但当与类似萼式的电导相结合时,它们可以在萼式被抑制的时期提高成功率,有效地在长时间快速刺激列车中支持可靠的输出。
微调传出信号的时序
非萼式输入最显著的影响体现在尖峰时序上。当真实的非萼式兴奋性突触后电位列车与模拟的萼式电导配对时,尖峰提前了数十微秒,在更高放电率和更强去极化下提前更多。使用简化模板,作者展示加入一个持续的兴奋性电导会使尖峰时序提前,而加入反向的抑制性电导则会使其延后。在测试范围内,联合的去极化与超极化驱动可将尖峰时序改变超过200微秒,并改变次间的时序抖动。这些时序位移恰好落在哺乳动物用于比较两只耳朵声到达时间以进行定位的时间范围内。 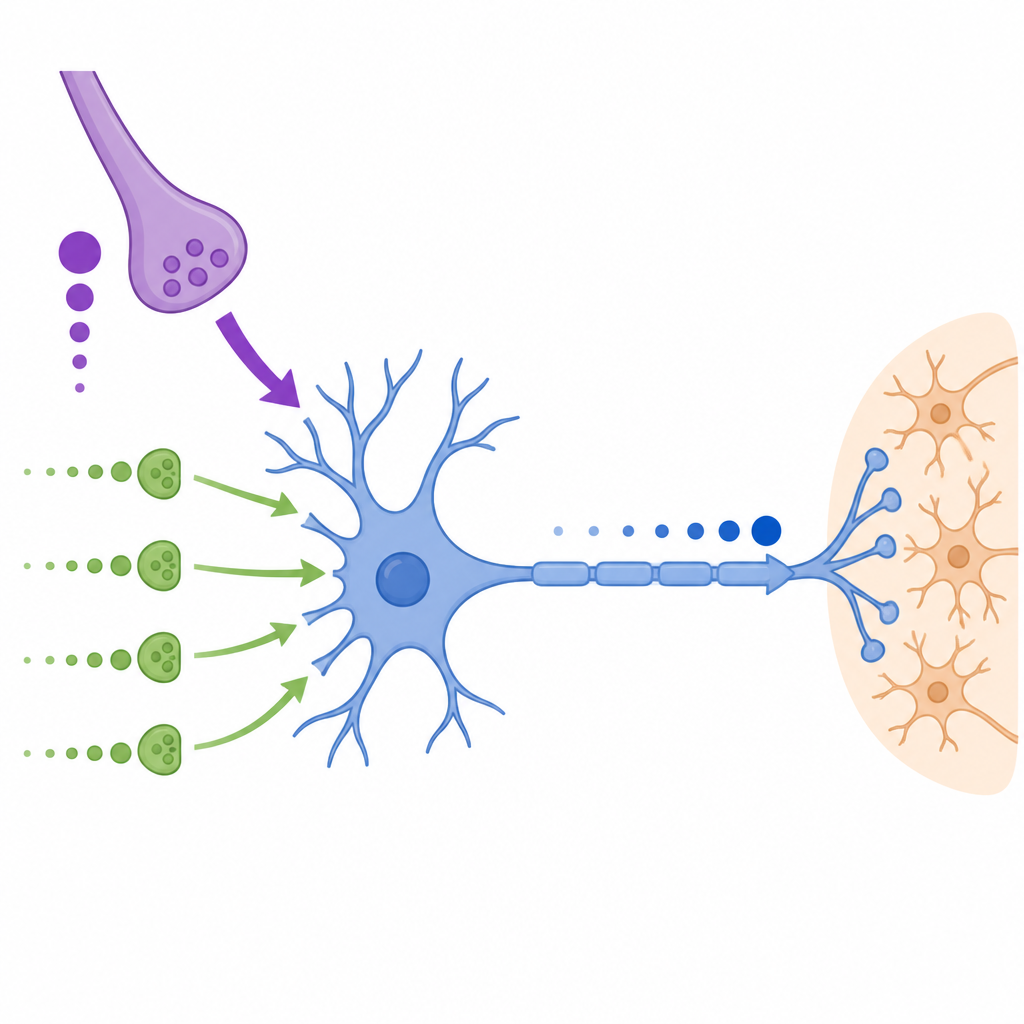
对真实听觉情境的意义
这项工作揭示MNTB不仅仅是被动复制其主输入的中继,而是一个由多重侧向输入雕塑发送到其他脑干中枢的抑制可靠性与时序的场所。通过提供可以是兴奋性或抑制性的缓慢变化背景驱动,非萼式输入“门控”尖峰的发生,使其提前或延后微小但具有行为意义的量。鉴于在沙鼠和厄特鲁斯坎鼩鼱中都发现了相似的突触模式,这种时序控制很可能反映了哺乳动物稳定与调整声源空间神经表征的一种通用策略。
引用: Console-Meyer, L., Jenzen, F., Kladisios, N. et al. Non-calyceal inputs gate the timing of calyx of Held evoked MNTB output. Commun Biol 9, 697 (2026). https://doi.org/10.1038/s42003-026-10321-w
关键词: 声音定位, 听觉脑干, 突触可塑性, 霍尔德萼式终板, 神经时序