Clear Sky Science · en
Non-calyceal inputs gate the timing of calyx of Held evoked MNTB output
How the brain keeps track of where sounds come from
Finding where a sound comes from, like a snapping twig in the dark, depends on the brain’s ability to send lightning-fast and precisely timed signals. This study looks at a tiny relay station in the auditory brainstem and shows that a set of overlooked side inputs can subtly retune the timing of outgoing signals, helping the brain build a reliable map of sound sources in space.
Two kinds of inputs to a key hearing relay
In the medial nucleus of the trapezoid body, a structure deep in the brainstem, principal neurons receive a giant main input called the calyx of Held as well as many smaller “non-calyceal” inputs on their soma and dendrites. The calyx delivers very large and fast signals but its strength drops during rapid, repeated activity. By contrast, the smaller inputs start out weak but become stronger when they are repeatedly activated and also keep releasing neurotransmitter in a more spread-out, asynchronous way. The researchers recorded these signals in brain slices from gerbils and Etruscan shrews and found this division of labor to be conserved across these distant mammalian species. 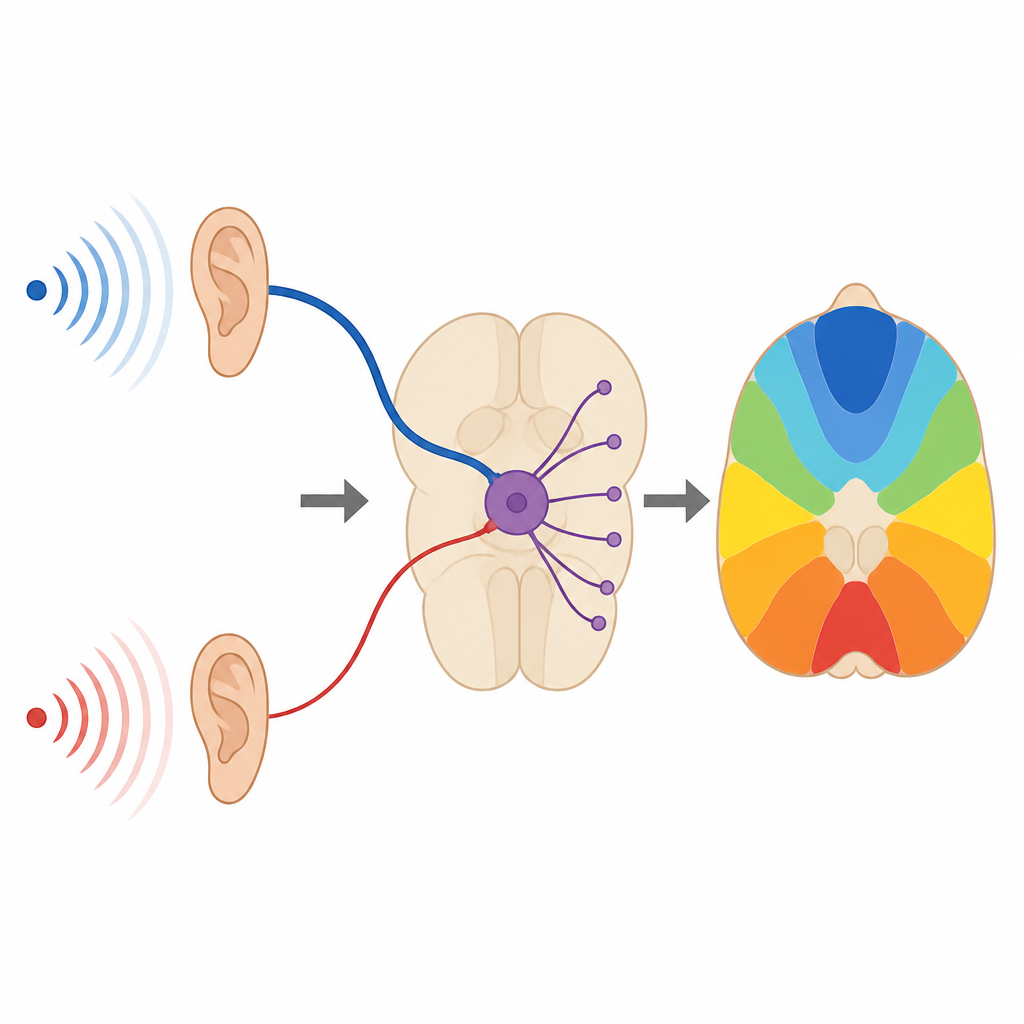
Short-term changes that shape ongoing activity
The team focused on short-term plasticity, the rapid strengthening or weakening of synapses during ongoing use. They showed that the calyx of Held strongly depresses during high-frequency stimulation, especially in gerbil, which means each successive response shrinks. Non-calyceal inputs, in contrast, show prominent facilitation, with responses that grow larger over the train and only mildly weaken at the very highest frequencies. These smaller inputs also produce delayed events that continue for hundreds of milliseconds after a stimulus train ends, effectively providing a steady depolarizing drive rather than sharply timed spikes. Raising calcium in the bath fluid converted the non-calyceal inputs from facilitating to depressing and revealed that their vesicle pools refill with time constants similar to those known for the calyx.
Simulating how combined inputs control spikes
To test how these different inputs work together, the authors used dynamic clamp, a technique that lets experimenters inject computer-generated conductances into real neurons. They constructed templates that mimicked the recorded behavior of the calyx and non-calyceal synapses, with and without a period of preconditioning that imitated ongoing background activity. When they applied these to MNTB neurons, they found that the probability, timing, and precision of action potentials mainly depended on the total synaptic conductance reaching the soma. The small inputs alone rarely drove spikes, but when added to the calyx-like conductance they could boost success rates during periods when the calyx was depressed, effectively supporting reliable output during long, fast stimulus trains.
Fine-tuning the timing of outgoing signals
The most striking effect of the non-calyceal inputs was on spike timing. When trains of real non-calyceal excitatory postsynaptic potentials were paired with simulated calyx conductances, spikes shifted earlier by tens of microseconds, with larger shifts at higher firing rates and stronger depolarization. Using simplified templates, the authors showed that adding a tonic excitatory conductance advanced spike timing, while an inverted, inhibitory version delayed it. Across the tested range, combined depolarizing and hyperpolarizing drives could shift spike timing by over 200 microseconds and also alter the trial-to-trial jitter. These shifts fall squarely within the time range used by mammals to compare sound arrival at the two ears for localization. 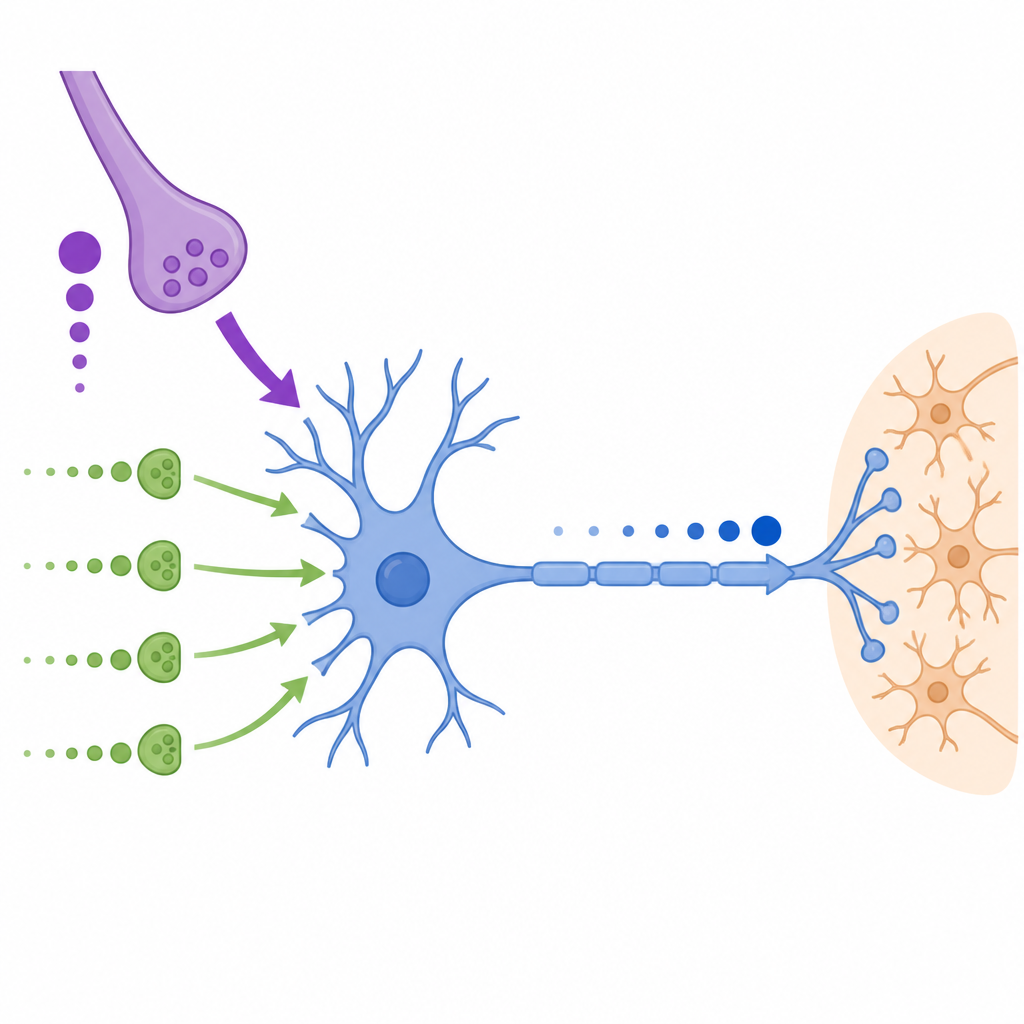
What this means for hearing in real life
This work reveals that the MNTB is not just a passive relay that copies its main input, but a site where multiple side inputs sculpt the reliability and timing of inhibition sent onward to other brainstem centers. By providing a slowly varying background drive that can be either excitatory or inhibitory, non-calyceal inputs “gate” when spikes occur, nudging them earlier or later by tiny but behaviorally meaningful amounts. Because similar synaptic patterns were found in both gerbil and Etruscan shrew, this timing control likely reflects a general strategy in mammals for stabilizing and adjusting the neural representation of sound sources in space.
Citation: Console-Meyer, L., Jenzen, F., Kladisios, N. et al. Non-calyceal inputs gate the timing of calyx of Held evoked MNTB output. Commun Biol 9, 697 (2026). https://doi.org/10.1038/s42003-026-10321-w
Keywords: sound localization, auditory brainstem, synaptic plasticity, calyx of Held, neural timing