Clear Sky Science · de
Nicht-kalyzale Eingänge steuern das Timing der vom Calyx of Held ausgelösten MNTB-Ausgabe
Wie das Gehirn nachvollzieht, woher Geräusche kommen
Die Lokalisierung einer Geräuschquelle, etwa das Knacken eines Zweigs im Dunkeln, beruht auf der Fähigkeit des Gehirns, blitzschnelle und präzise getaktete Signale zu liefern. Diese Studie betrachtet eine winzige Schaltstelle im auditorischen Hirnstamm und zeigt, dass eine Reihe bislang wenig beachteter seitlicher Eingänge das Timing der ausgehenden Signale subtil nachstimmen kann, wodurch dem Gehirn geholfen wird, eine verlässliche Karte der Schallquellen im Raum zu erstellen.
Zwei Arten von Eingängen zu einem wichtigen Hörrelais
Im medialen Kern des Trapezkörpers, einer tief im Hirnstamm gelegenen Struktur, erhalten Hauptneurone einen riesigen Haupteingang, den Calyx of Held, sowie zahlreiche kleinere „nicht-kalyzale“ Eingänge an Soma und Dendriten. Der Calyx liefert sehr große und schnelle Signale, deren Stärke bei rascher, wiederholter Aktivität jedoch abnimmt. Im Gegensatz dazu sind die kleineren Eingänge anfangs schwach, verstärken sich aber bei wiederholter Aktivierung und setzen Neurotransmitter zudem in einem stärker asynchronen, über einen längeren Zeitraum verteilten Muster frei. Die Forscher zeichneten diese Signale in Hirnschnitten von Gerbils und etruskischen Spitzmäusen auf und fanden diese Arbeitsteilung über diese entfernten Säugetierarten hinweg konserviert. 
Kurzfristige Veränderungen, die die laufende Aktivität formen
Das Team konzentrierte sich auf kurzfristige Plastizität, also die schnelle Verstärkung oder Abschwächung von Synapsen während laufender Nutzung. Sie zeigten, dass der Calyx of Held bei hochfrequenter Stimulation stark deprimiert, insbesondere beim Gerbil, wodurch jede aufeinanderfolgende Antwort kleiner wird. Nicht-kalyzale Eingänge dagegen zeigen ausgeprägte Facilitation: die Antworten wachsen im Verlauf eines Reizträgers und schwächen sich nur bei den höchsten Frequenzen leicht ab. Diese kleineren Eingänge erzeugen außerdem verzögerte Ereignisse, die noch Hunderte von Millisekunden nach Ende eines Reiztrains andauern und damit eher einen stetigen depolarisierenden Antrieb bereitstellen als scharf getaktete Spikes. Eine Erhöhung des Calciumgehalts im Badmedium wandelte die nicht-kalyzalen Eingänge von facilitierend zu deprimierend und zeigte, dass sich deren Vesikelvorräte mit Zeitkonstanten auffüllen, die denen des Calyx ähneln.
Simulation, wie kombinierte Eingänge Spikes steuern
Um zu testen, wie diese unterschiedlichen Eingänge zusammenwirken, verwendeten die Autoren Dynamic Clamp, eine Technik, mit der experimentell computergenerierte Leitfähigkeiten in echte Neurone injiziert werden können. Sie konstruierten Vorlagen, die das aufgezeichnete Verhalten des Calyx und der nicht-kalyzalen Synapsen nachahmten, mit und ohne eine Periode der Vorkonditionierung, die andauernde Hintergrundaktivität imitierte. Als sie diese auf MNTB-Neurone anwandten, stellten sie fest, dass die Wahrscheinlichkeit, das Timing und die Präzision von Aktionspotenzialen hauptsächlich von der gesamten synaptischen Leitfähigkeit abhängen, die das Soma erreicht. Die kleinen Eingänge allein trieben selten Spikes, aber in Kombination mit einer calyx-ähnlichen Leitfähigkeit konnten sie die Erfolgsraten in Phasen steigern, in denen der Calyx deprimiert war, und so während langer, schneller Stimulusreihen eine verlässliche Ausgabe unterstützen.
Feinabstimmung des Timings ausgehender Signale
Die auffälligste Wirkung der nicht-kalyzalen Eingänge zeigte sich im Spike-Timing. Wenn Reihen realer nicht-kalyzaler exzitatorischer postsynaptischer Potentiale mit simulierten Calyx-Leitfähigkeiten gepaart wurden, verschoben sich Spikes um einige zehn Mikrosekunden nach vorn, mit größeren Verschiebungen bei höheren Feuerraten und stärkerer Depolarisation. Mit vereinfachten Vorlagen zeigten die Autoren, dass das Hinzufügen einer tonischen exzitatorischen Leitfähigkeit das Spike-Timing vorverlegte, während eine invertierte, inhibitorische Version es verzögerte. Innerhalb des getesteten Bereichs konnten kombinierte depolarisierende und hyperpolarisierende Antriebe das Spike-Timing um mehr als 200 Mikrosekunden verschieben und auch die Trial-zu-Trial-Jitter verändern. Diese Verschiebungen liegen genau im Zeitbereich, den Säugetiere zur Vergleichung der Schalleinfallzeiten an beiden Ohren für die Lokalisation nutzen. 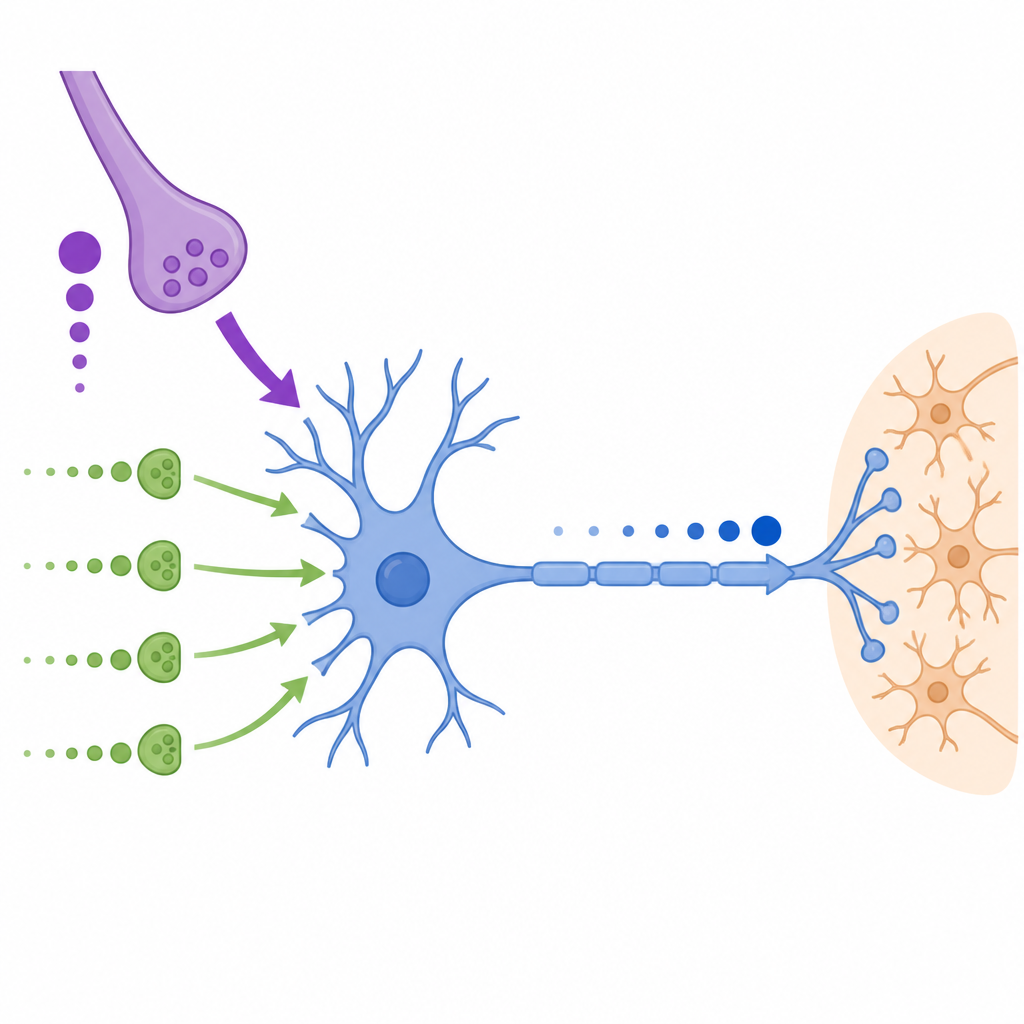
Was das für das Hören im echten Leben bedeutet
Diese Arbeit zeigt, dass die MNTB nicht nur ein passives Relais ist, das seinen Haupteingang kopiert, sondern ein Ort, an dem multiple seitliche Eingänge die Zuverlässigkeit und das Timing der Hemmung formen, die an andere Hirnstammzentren weitergeleitet wird. Indem sie einen langsam variierenden Hintergrundantrieb bereitstellen können, der exzitatorisch oder inhibitorisch sein kann, „steuern“ nicht-kalyzale Eingänge, wann Spikes auftreten, und schubsen sie um kleine, aber verhaltensrelevante Zeiträume früher oder später. Da ähnliche synaptische Muster sowohl beim Gerbil als auch bei der etruskischen Spitzmaus gefunden wurden, spiegelt diese Timing-Kontrolle wahrscheinlich eine allgemeine Strategie von Säugetieren wider, die neuronale Repräsentation von Schallquellen im Raum zu stabilisieren und anzupassen.
Zitation: Console-Meyer, L., Jenzen, F., Kladisios, N. et al. Non-calyceal inputs gate the timing of calyx of Held evoked MNTB output. Commun Biol 9, 697 (2026). https://doi.org/10.1038/s42003-026-10321-w
Schlüsselwörter: Schallortung, auditorischer Hirnstamm, synaptische Plastizität, Calyx of Held, neurales Timing