Clear Sky Science · it
Ingressi non caliciformi regolano il timing dell’output MNTB evocato dal calyx of Held
Come il cervello tiene traccia della provenienza dei suoni
Trovare da dove proviene un suono, come il crepitio di un ramo nel buio, dipende dalla capacità del cervello di inviare segnali fulminei e temporizzati con precisione. Questo studio esamina una piccola stazione di relè nel tronco encefalico uditivo e mostra che un insieme di ingressi laterali finora trascurati può ritunare sottilmente il timing dei segnali in uscita, aiutando il cervello a costruire una mappa affidabile delle sorgenti sonore nello spazio.
Due tipi di ingressi a un relè chiave per l’udito
Nel nucleo mediale del corpo trapezoide, una struttura profonda nel tronco encefalico, i neuroni principali ricevono un grande ingresso principale chiamato calyx of Held e molti ingressi più piccoli “non caliciformi” sul soma e sui dendriti. Il calyx fornisce segnali molto grandi e veloci ma la sua efficacia diminuisce durante attività rapida e ripetuta. Al contrario, gli ingressi più piccoli partono deboli ma si rafforzano quando vengono attivati ripetutamente e rilasciano anche neurotrasmettitore in modo più diffuso e asincrono. I ricercatori hanno registrato questi segnali in fette di cervello di gerbilli e musarole etrusche e hanno trovato che questa divisione di compiti è conservata in queste specie di mammiferi distanti. 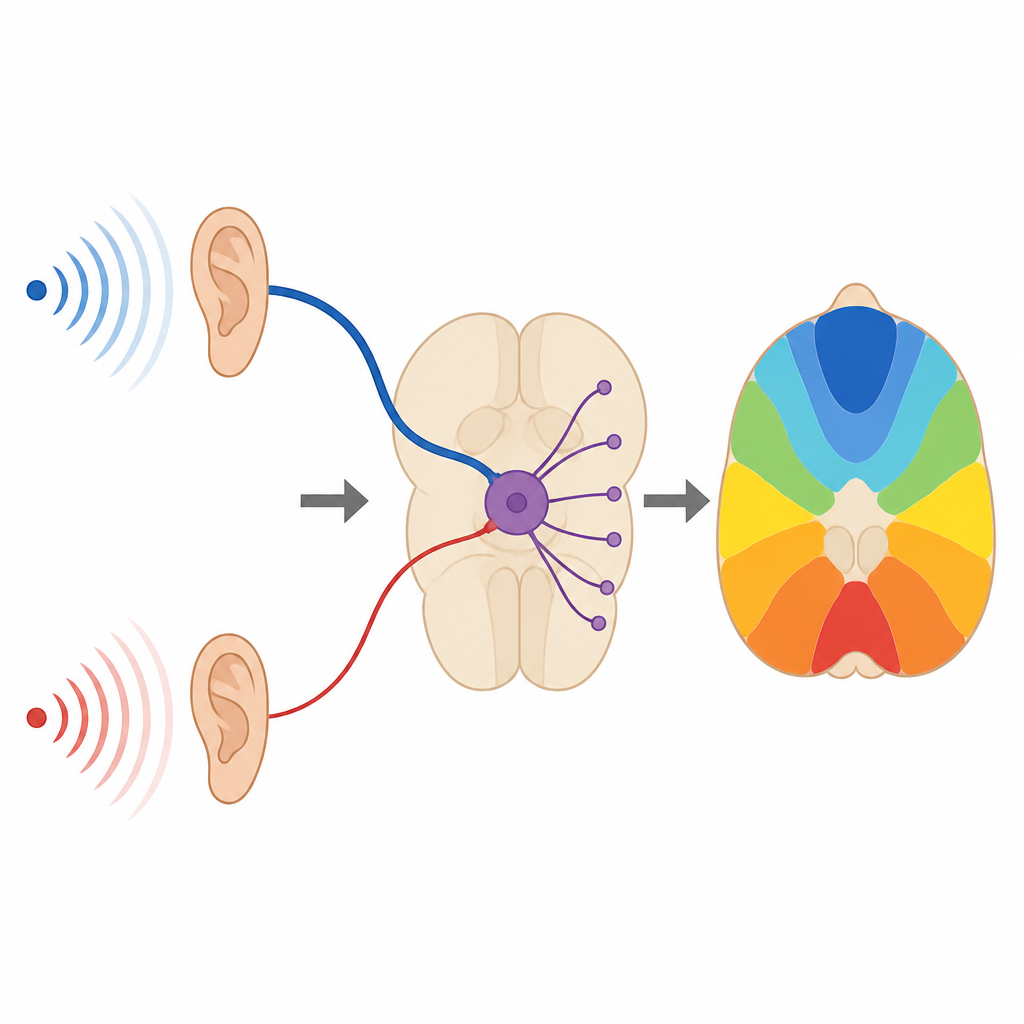
Cambiamenti a breve termine che modellano l’attività in corso
Il gruppo si è focalizzato sulla plasticità a breve termine, l’indebolimento o il rafforzamento rapido delle sinapsi durante l’uso continuativo. Hanno mostrato che il calyx of Held si depotenzia fortemente durante stimolazione ad alta frequenza, specialmente nel gerbillo, il che significa che ogni risposta successiva si riduce. Gli ingressi non caliciformi, al contrario, mostrano una marcata facilitazione, con risposte che crescono lungo la serie e solo un lieve indebolimento alle frequenze più elevate. Questi ingressi più piccoli producono anche eventi ritardati che persistono per centinaia di millisecondi dopo la fine di una serie di stimoli, fornendo di fatto una spinta depolarizzante costante piuttosto che spike temporizzati in modo netto. Aumentare il calcio nel bagno sperimentale convertiva gli ingressi non caliciformi da facilitanti a depressivi e rivelava che i loro pool di vescicole si ricaricano con costanti di tempo simili a quelle note per il calyx.
Simulare come gli ingressi combinati controllano gli spike
Per testare come questi diversi ingressi lavorino insieme, gli autori hanno usato il dynamic clamp, una tecnica che permette di iniettare conduttanze generate al computer in neuroni reali. Hanno costruito template che imitavano il comportamento registrato delle sinapsi caliciformi e non caliciformi, con e senza un periodo di precondizionamento che imitava l’attività di fondo continua. Applicandoli ai neuroni MNTB, hanno riscontrato che la probabilità, il timing e la precisione dei potenziali d’azione dipendono principalmente dalla conduttanza sinaptica totale che raggiunge il soma. I piccoli ingressi da soli raramente generavano spike, ma quando aggiunti alla conduttanza simile al calyx potevano aumentare i tassi di successo durante i periodi in cui il calyx era depresso, supportando di fatto un’uscita affidabile durante serie di stimoli lunghe e rapide.
Aggiustare finemente il timing dei segnali in uscita
L’effetto più sorprendente degli ingressi non caliciformi riguardava il timing degli spike. Quando treni di potenziali postsinaptici eccitatori non caliciformi reali venivano abbinati a conduttanze calyx simulate, gli spike si spostavano in anticipo di decine di microsecondi, con spostamenti maggiori a tassi di scarica più elevati e con depolarizzazione più forte. Usando template semplificati, gli autori hanno mostrato che aggiungere una conduttanza eccitatoria tonica anticipava il timing degli spike, mentre una versione invertita, inibitoria, lo ritardava. Nell’intervallo testato, spinte depolarizzanti e iperpolarizzanti combinate potevano spostare il timing degli spike di oltre 200 microsecondi e alterare anche la variabilità trial-to-trial. Questi spostamenti ricadono nettamente nell’intervallo temporale che i mammiferi usano per confrontare l’arrivo del suono alle due orecchie per la localizzazione. 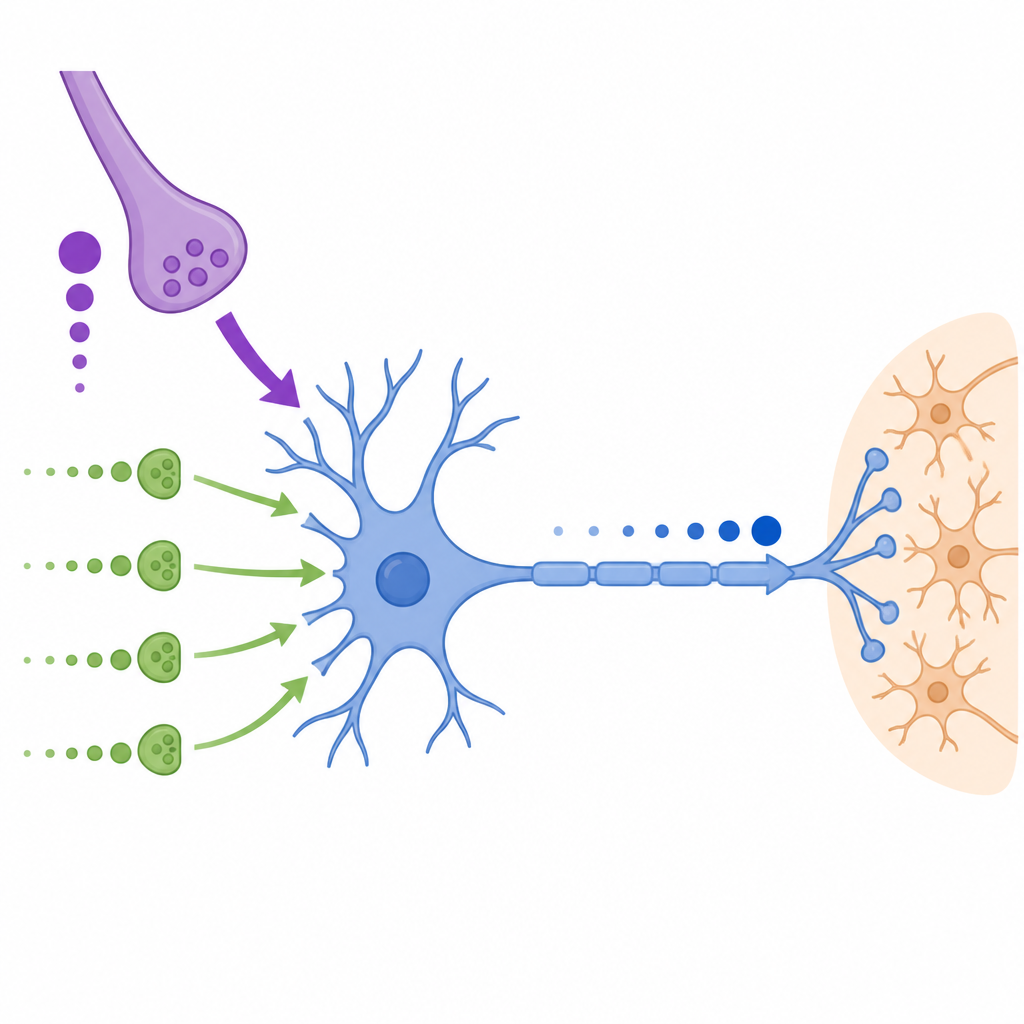
Cosa significa questo per l’udito nella vita reale
Questo lavoro rivela che l’MNTB non è solo un relè passivo che copia il suo ingresso principale, ma un sito in cui molteplici ingressi laterali scolpiscono l’affidabilità e il timing dell’inibizione inviata ad altri centri del tronco encefalico. Fornendo una spinta di fondo a variazione lenta che può essere eccitatoria o inibitoria, gli ingressi non caliciformi “regolano” quando si verificano gli spike, spostandoli in anticipo o in ritardo di quantità piccole ma comportamentalmente rilevanti. Poiché schemi sinaptici simili sono stati trovati sia nel gerbillo che nella musarola etrusca, questo controllo del timing probabilmente riflette una strategia generale nei mammiferi per stabilizzare e aggiustare la rappresentazione neurale delle sorgenti sonore nello spazio.
Citazione: Console-Meyer, L., Jenzen, F., Kladisios, N. et al. Non-calyceal inputs gate the timing of calyx of Held evoked MNTB output. Commun Biol 9, 697 (2026). https://doi.org/10.1038/s42003-026-10321-w
Parole chiave: localizzazione del suono, tronco encefalico uditivo, plasticità sinaptica, calyx of Held, tempistica neuronale