Clear Sky Science · zh
Runx 蛋白的磷酸化控制辅助 CD4+ T 细胞与细胞毒性 CD8+ T 细胞谱系的选择
免疫细胞如何决定它们的职责
我们的免疫系统依赖于两类 T 细胞:一种协调防御(辅助 CD4+ 细胞),另一种直接杀死被感染或癌变的细胞(细胞毒性 CD8+ 细胞)。本文提出了一个看似简单的问题:当幼稚的 T 细胞在胸腺中成熟时,它们如何决定走上这两种截然不同的职业道路?作者发现了 Runx 蛋白上单个氨基酸上的微小化学标记,作为一个分子开关,将外部信号与这一决定命运的选择连接起来。
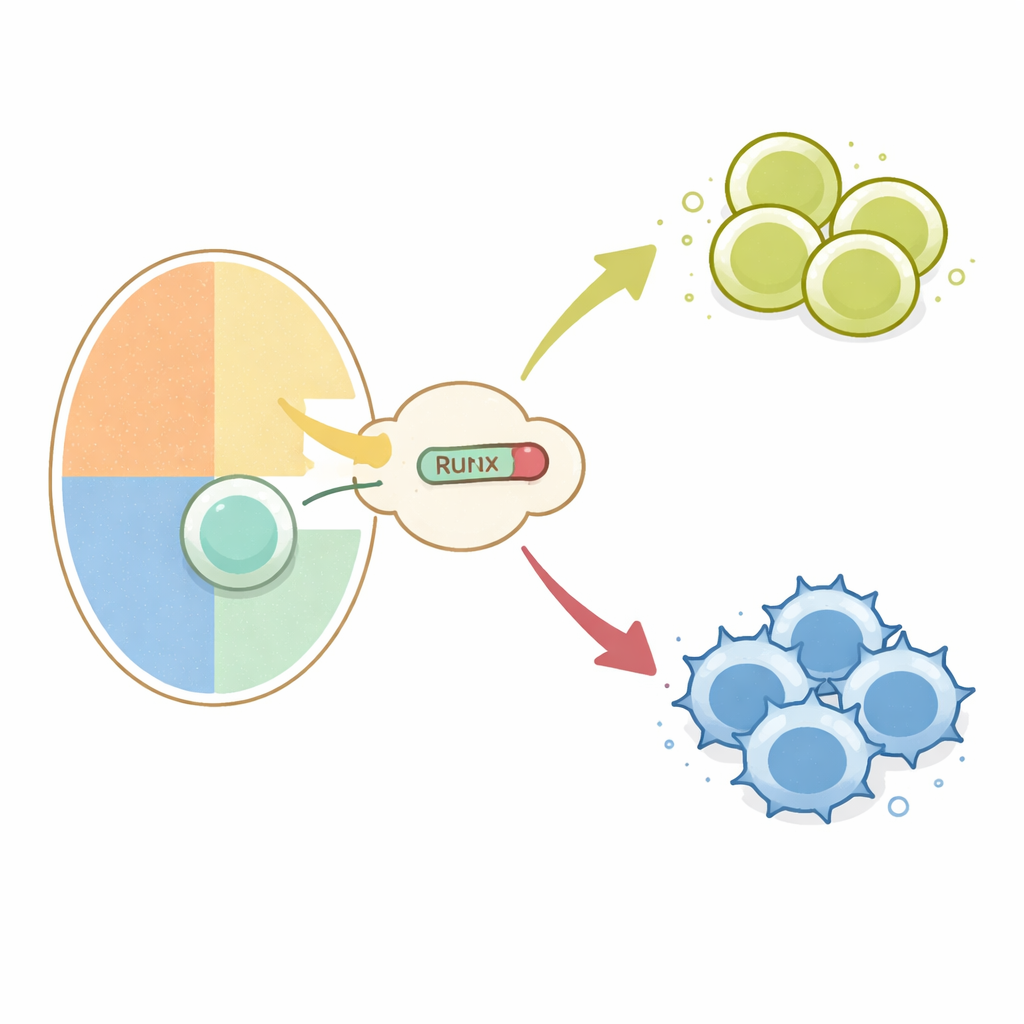
胸腺内的一个分岔路口
所有常规 T 细胞都起始于胸腺中的未成熟前体,它们在细胞表面同时呈现 CD4 和 CD8 分子。在检验受体与自身分子相互作用的过程中,这些细胞被指引为辅助(CD4)或杀伤(CD8)T 细胞。早期研究表明,关闭两个基因 Cd4 和 Thpok 对于走向杀伤命运至关重要。这种关闭由 Runx 家族转录因子与 TLE 共抑制蛋白在称为沉默子(silencer)的特定 DNA 区域共同驱动。未知之处在于,如何将区分与主要组织相容性复合体 I 类(导致杀手命运)与 II 类(导致辅助命运)相互作用的 T 细胞受体信号,接入这一 Runx–TLE 的沉默机制。
单个字母的改变将 Runx 变成制动器
Runx 蛋白以一个短序列 WRPY 结尾,可与 TLE 蛋白对接。研究组构建了小鼠模型,将末端氨基酸酪氨酸(Y)替换为色氨酸(W)或苯丙氨酸(F),生成名为 RunxWRPW 或 RunxWRPF 的变体。这些细微改变将 Runx1 和 Runx3 转变为强大且几乎始终处于“开关关闭”状态的抑制因子。表达这些突变形式的鼠类在许多依赖 Runx 的细胞类型上出现严重缺陷:CD8 T 细胞几乎缺失,自然杀伤细胞和某些先天淋巴样细胞无法发育,而在 Runx1 的情况下,胚胎因失去造血干细胞而死亡。这表明正常的末端 Y 残基对于维持 Runx 在激活和抑制基因之间的平衡是必需的。
将辅助细胞改写为杀手细胞
为聚焦于 T 细胞命运选择,作者利用遗传开关在发育中的胸腺细胞中特异性地启动正常 Runx 或 RunxWRPW/RunxWRPF。当突变 Runx 从双阳性阶段起表达时,原本识别 II 类分子并将成为 CD4 辅助细胞的细胞,反而被重定向进入 CD8 谱系并失去 CD4 表达。即使携带已知能强制辅助命运的转基因受体的 T 细胞,也被转变为类似杀手的细胞。这种重定向依赖于 Cd4 和 Thpok 基因中的相同沉默子,以及 TLE 蛋白,表明过强的 Runx–TLE 抑制能够覆盖受体特异性与谱系之间通常的联系。
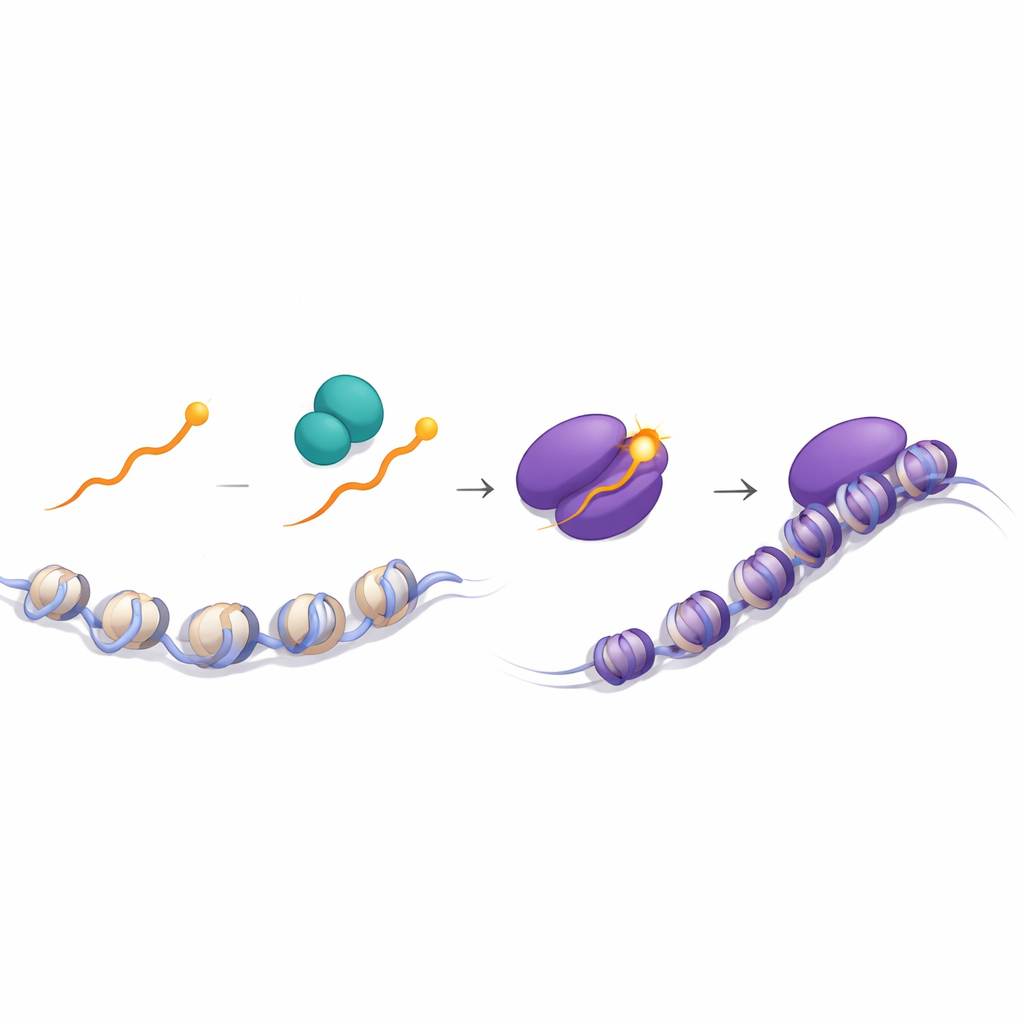
感知外部线索的磷酸化开关
关键线索来自对正常 Runx1 尾端的分析。利用质谱,作者发现末端的 Y 在 CD8 胸腺细胞中的磷酸化水平远高于 CD4 胸腺细胞。这一修饰显著增强了 Runx1 与 TLE3 的相互作用。在缺失酪氨酸激酶 Lck 的细胞中,这种磷酸化几乎不存在,指向作为 T 细胞受体信号中心传导子 Lck 及其伙伴 Zap70 的作用。成像实验证明,Runx1 主要在细胞质中与 Lck 和 Zap70 相遇,并且在通过 I 类分子被信号刺激的细胞中这种相遇更为频繁。结构模拟支持这样一种观点:磷酸化的 Y,或者被 F 或 W 替代的 Y,会稳定 Runx 与 TLE 的 WD40 结构域结合,促进在沉默子上组装成强效的抑制复合体。
从微小化学标记到免疫细胞身份
综上,研究提出 Runx 蛋白末端单个酪氨酸的磷酸化充当一个灵敏的调节旋钮:在受到 I 类信号的胸腺细胞中,较高的磷酸化更有效地招募 TLE 到 Cd4 和 Thpok 基因的沉默子,关闭它们并锁定细胞毒性 CD8 命运;在受到 II 类信号的细胞中,低磷酸化使 Runx 与 TLE 的结合较松,沉默子失效,辅助 CD4 程序得以进行。对非专业读者来说,这一结论表明免疫系统可以使用位于蛋白一端的极其微小化学修饰,将细胞受体传入的细微信号翻译为在两类核心 T 细胞之间的全或无选择,体现了微调分子开关如何支撑发育过程中稳健决策的精密性。
引用: Ogawa, C., Okuyama, K., Kojo, S. et al. Phosphorylation of Runx protein controls helper CD4+ T cell versus cytotoxic CD8+ T cell lineage choice. Nat Immunol 27, 799–811 (2026). https://doi.org/10.1038/s41590-026-02441-6
关键词: T 细胞分化, Runx 磷酸化, CD4 与 CD8 谱系, 基因沉默, 胸腺发育