Clear Sky Science · de
Phosphorylierung des Runx-Proteins steuert die Wahl zwischen Helfer-CD4+- und zytotoxischer CD8+-T‑Zell‑Linie
Wie Immunzellen ihre Aufgabe bestimmen
Unser Immunsystem beruht auf T‑Zellen, die entweder die Abwehr koordinieren (Helfer‑CD4+-Zellen) oder infizierte und entartete Zellen direkt abtöten (zytotoxische CD8+-Zellen). Diese Arbeit stellt eine auf den ersten Blick einfache Frage: Wenn junge T‑Zellen im Thymus heranreifen, wie treffen sie die Entscheidung für eine von zwei so unterschiedlichen Karrieren? Die Autoren entdeckten eine winzige chemische Markierung an einer einzigen Aminosäure eines Proteins namens Runx, die als molekularer Schalter wirkt und Außensignale mit dieser lebensbestimmenden Wahl verbindet.
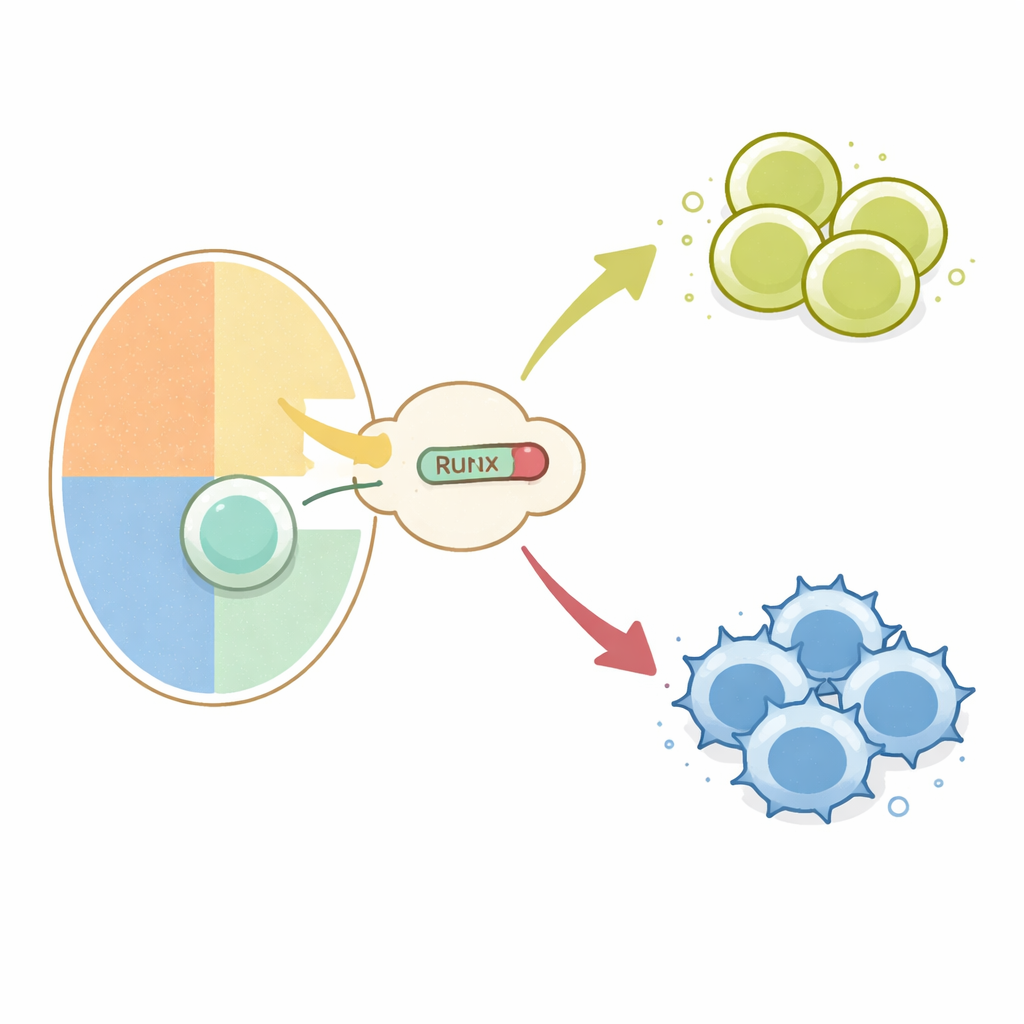
Eine Weggabelung im Thymus
Alle konventionellen T‑Zellen beginnen als unreife Vorläufer im Thymus und zeigen sowohl CD4‑ als auch CD8‑Oberflächenmoleküle. Beim Testen ihrer Rezeptoren an körpereigenen Molekülen werden sie angewiesen, entweder Helfer (CD4) oder Killer (CD8) zu werden. Frühere Arbeiten zeigten, dass das Abschalten zweier Gene, Cd4 und Thpok, für das Killer‑Schicksal essenziell ist. Diese Abschaltung wird von Runx‑Familien‑Transkriptionsfaktoren in Zusammenarbeit mit TLE‑Kernrepressoren an speziellen DNA‑Regionen, sogenannten Silencern, vorangetrieben. Unbekannt war, wie Signale vom T‑Zellrezeptor, die Interaktionen mit MHC Klasse I (führt zu Killern) versus Klasse II (führt zu Helfern) unterscheiden, in diese Runx–TLE‑Stillschaltmaschinerie eingespeist werden.
Eine Ein‑Buchstaben‑Änderung macht Runx zum Bremsen
Die Runx‑Proteine enden in einem kurzen Motiv, WRPY, das an TLE‑Proteine andockt. Das Team erzeugte Mäuse, bei denen die letzte Aminosäure, Tyrosin (Y), durch Tryptophan (W) oder Phenylalanin (F) ersetzt wurde und damit Versionen entstanden, die als RunxWRPW bzw. RunxWRPF bezeichnet wurden. Diese subtilen Veränderungen verwandelten Runx1 und Runx3 in starke, nahezu immer aktive Repressoren. Mäuse, die diese mutierten Formen exprimierten, zeigten schwere Defekte in vielen Runx‑abhängigen Zelltypen: CD8‑T‑Zellen waren nahezu abwesend, natürliche Killerzellen und bestimmte angeborene lymphoide Zellen entwickelten sich nicht, und im Fall von Runx1 führten die Veränderungen zum Embryotod mit Verlust blutbildender Stammzellen. Das zeigte, dass die normale terminale Y‑Restgruppe nötig ist, um Runx im Gleichgewicht zwischen Aktivierung und Repression von Genen zu halten.
Umlenkung von Helfern zu Killern
Um sich auf die T‑Zell‑Schicksalswahl zu konzentrieren, nutzten die Autoren einen genetischen Schalter, um entweder normales Runx oder RunxWRPW/RunxWRPF spezifisch in sich entwickelnden Thymozyten anzuschalten. Wenn das mutierte Runx ab dem Doppel‑positiv‑Stadium exprimiert wurde, wurden Zellen, die normalerweise Klasse‑II‑Moleküle erkennen und zu CD4‑Helfern würden, stattdessen in die CD8‑Linie umgelenkt und verloren die CD4‑Expression. Sogar T‑Zellen mit einem transgenen Rezeptor, der normalerweise das Helfer‑Schicksal erzwingt, wurden in Killer‑ähnliche Zellen umgewandelt. Diese Umlenkung hing von denselben Silencern in den Cd4‑ und Thpok‑Genen und von TLE‑Proteinen ab, was zeigt, dass überstarke Runx–TLE‑Repression die übliche Verbindung zwischen Rezeptorspezifität und Linie außer Kraft setzen kann.
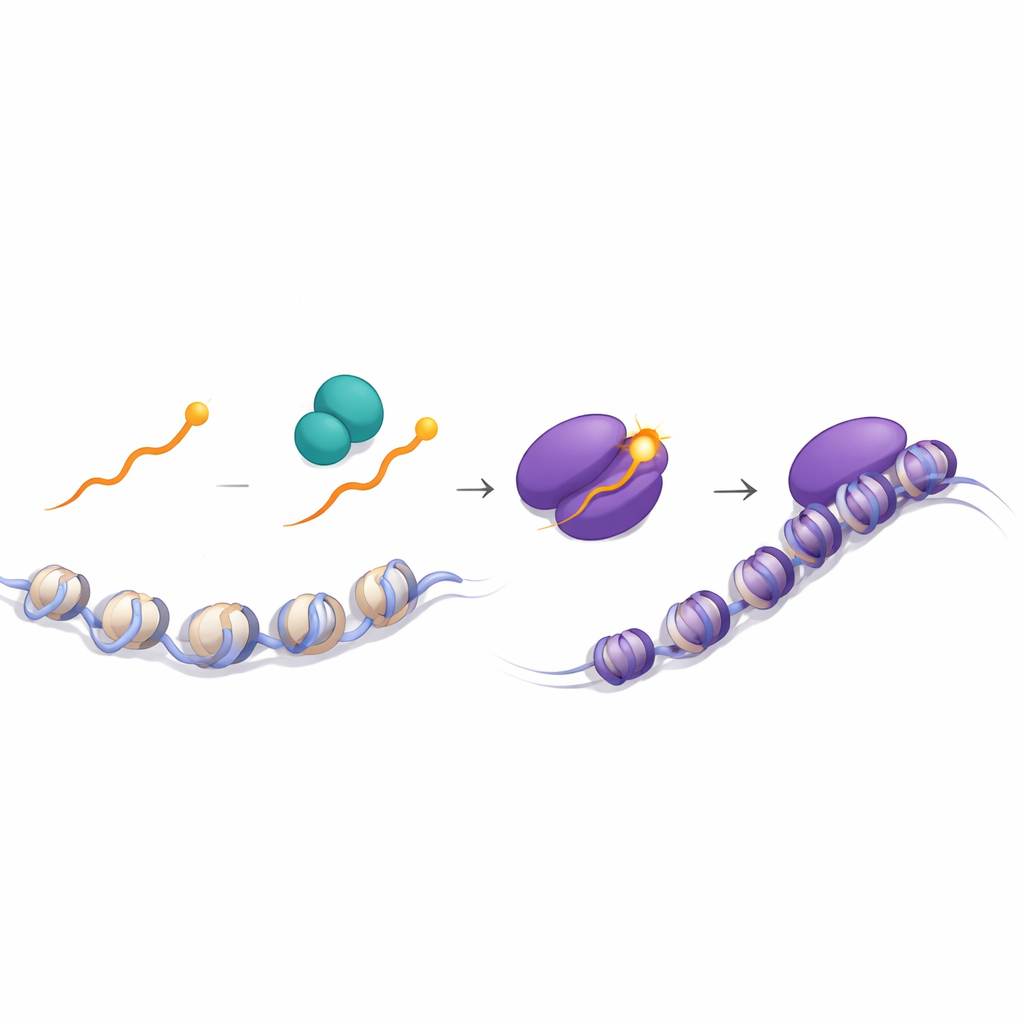
Ein Phosphorylierungs‑Schalter, der Außensignale erkennt
Die entscheidende Einsicht ergab sich aus der Untersuchung des normalen Runx1‑Schwanzes. Mithilfe von Massenspektrometrie fanden die Autoren heraus, dass das terminale Y in CD8‑Thymozyten deutlich stärker phosphoryliert ist — mit einer Phosphatgruppe versehen — als in CD4‑Thymozyten. Diese Modifikation stärkt die Wechselwirkung von Runx1 mit TLE3 erheblich. In Zellen, denen die Tyrosinkinase Lck fehlte, war diese Phosphorylierung nahezu nicht vorhanden, was Lck und dessen Partner Zap70, beides zentrale Überträger von T‑Zellrezeptorsignalen, involvierte. Bildgebende Experimente zeigten, dass Runx1 Lck und Zap70 vorwiegend im Zytoplasma begegnet und dass solche Begegnungen in Zellen, die über Klasse‑I‑Moleküle signalisiert werden, häufiger sind. Strukturelle Simulationen stützten die Idee, dass ein phosphoryliertes Y — oder ein durch F oder W ersetztes Y — die Bindung von Runx an die WD40‑Domäne von TLE stabilisiert und den Zusammenbau eines potenten Repressionskomplexes an Silencern fördert.
Von der winzigen chemischen Markierung zur Identität der Immunzelle
Insgesamt schlagen die Autoren vor, dass die Phosphorylierung der einzelnen terminalen Tyrosin‑Restgruppe an Runx‑Proteinen als empfindlicher Drehknopf dient: In Klasse‑I‑signalierten Thymozyten führt die höhere Phosphorylierung zu einer effizienteren Rekrutierung von TLE an Silencer in den Cd4‑ und Thpok‑Genen, wodurch diese abgeschaltet und das zytotoxische CD8‑Schicksal festgelegt wird. In Klasse‑II‑signalierten Zellen bleibt die Phosphorylierung gering, Runx bindet weniger fest an TLE, die Silencer sind unwirksam und das Helfer‑CD4‑Programm schreitet fort. Für eine breite Leserschaft lautet die Botschaft: Das Immunsystem kann eine winzige chemische Modifikation an einem Ende eines Proteins nutzen, um subtile Rezeptorsignale in eine Alles‑oder‑Nichts‑Entscheidung zwischen zwei zentralen T‑Zell‑Typen zu übersetzen — ein Beispiel dafür, wie fein abgestimmte molekulare Schalter robuste Entwicklungsentscheidungen ermöglichen.
Zitation: Ogawa, C., Okuyama, K., Kojo, S. et al. Phosphorylation of Runx protein controls helper CD4+ T cell versus cytotoxic CD8+ T cell lineage choice. Nat Immunol 27, 799–811 (2026). https://doi.org/10.1038/s41590-026-02441-6
Schlüsselwörter: T‑Zell‑Differenzierung, Runx‑Phosphorylierung, CD4 versus CD8 Linie, Genstilllegung, thymische Entwicklung