Clear Sky Science · ru
Фосфорилирование белка Runx контролирует выбор линейности между хелперными CD4+ и цитотоксическими CD8+ T‑клетками
Как иммунные клетки выбирают свою роль
Наша иммунная система опирается на T‑клетки, которые либо координируют защиту (хелперные CD4+ клетки), либо напрямую уничтожают инфицированные и опухолевые клетки (цитотоксические CD8+ клетки). В этой статье поставлен на первый взгляд простой вопрос: когда молодые T‑клетки созревают в тимусе, как они решают, какую из этих двух кардинально разных профессий выбрать? Авторы выявляют крошечную химическую метку на одной аминокислоте белка Runx, которая действует как молекулярный переключатель, связывая внешние сигналы с этим жизненно важным выбором.
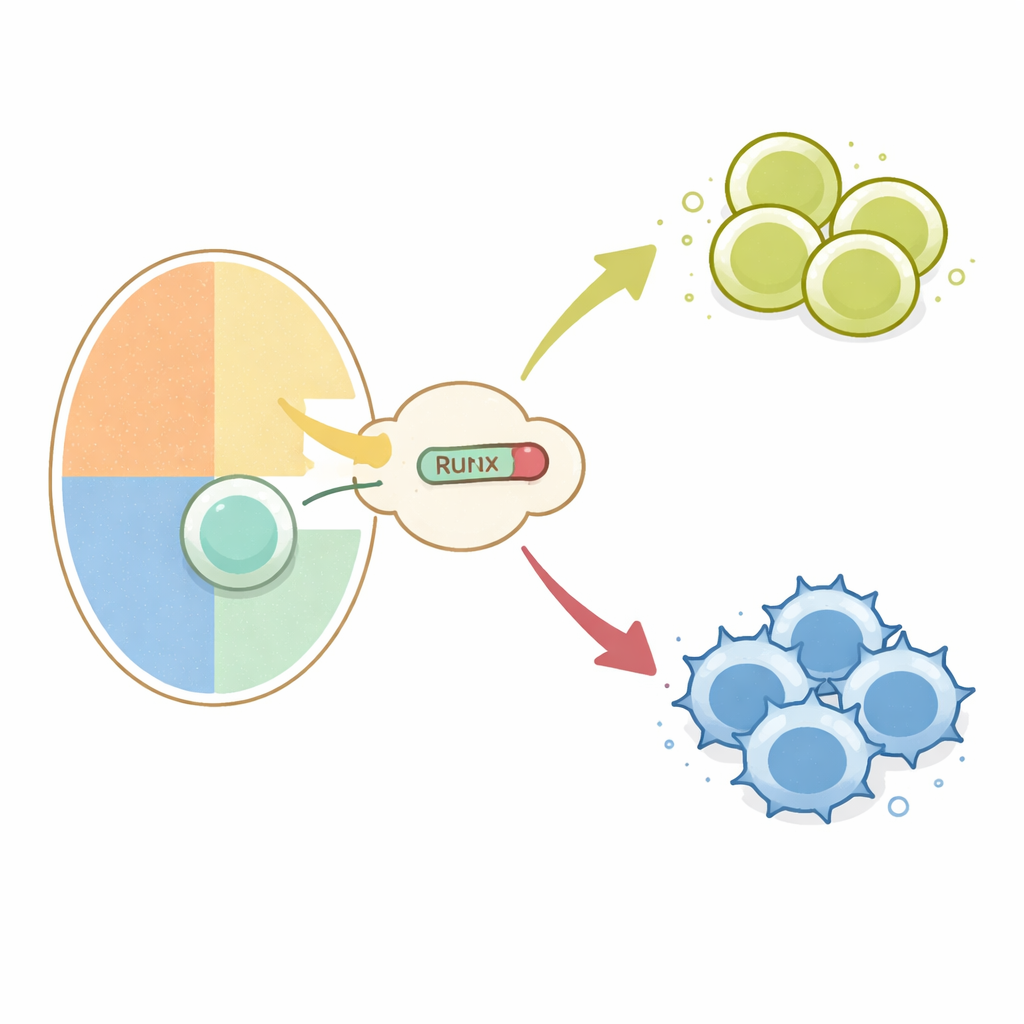
Перекрёсток внутри тимуса
Все обычные T‑клетки начинаются как незрелые предшественники в тимусе, которые экспрессируют на поверхности и CD4, и CD8 молекулы. При проверке своих рецепторов на самостные молекулы им даётся инструкция стать либо хелперами (CD4), либо киллерами (CD8). Ранние исследования показали, что выключение двух генов, Cd4 и Thpok, необходимо для судьбы киллера. Это подавление обеспечивается транскрипционными факторами семейства Runx в сотрудничестве с корепрессорами TLE на специальных участках ДНК, называемых сайленсерами. Не было ясно, как сигналы от T‑клеточного рецептора, которые отличают взаимодействия с молекулами главного комплекса гистосовместимости класса I (ведущие к киллерам) и класса II (ведущие к хелперам), внедряются в эту систему подавления Runx–TLE.
Однобуквенное изменение превращает Runx в тормоз
Белки Runx заканчиваются коротким мотивом WRPY, который связывается с белками TLE. Команда создала мышей, в которых последняя аминокислота, тирозин (Y), была заменена на триптофан (W) или фенилаланин (F), получив варианты, обозначенные RunxWRPW или RunxWRPF. Эти тонкие изменения превратили Runx1 и Runx3 в мощные, почти постоянно включённые репрессоры. У мышей, экспрессирующих эти мутантные формы, наблюдались серьёзные дефекты во многих типах клеток, зависящих от Runx: CD8‑клеток почти не было, естественные киллеры и некоторые врождённые лимфоидные клетки не развивались, а при мутации Runx1 эмбрионы погибали с потерей гемопоэтических стволовых клеток. Это показало, что нормальный терминальный остаток Y необходим для поддержания баланса Runx между активацией и репрессией генов.
Перепрограммирование хелперов в киллеры
Чтобы сфокусироваться на выборе судьбы T‑клеток, авторы использовали генетический переключатель для включения либо нормального Runx, либо RunxWRPW/RunxWRPF специально в развивающихся тимоцитах. Когда мутантный Runx экспрессировался с этапа двойнопозитивных клеток и далее, клетки, которые обычно распознают молекулы класса II и должны были стать CD4‑хелперами, вместо этого перенаправлялись в линию CD8 и теряли экспрессию CD4. Даже T‑клетки с трансгенным рецептором, который обычно навязывает хелперную судьбу, были превращены в клетки, похожие на киллеров. Это перенаправление зависело от тех же сайленсеров в генах Cd4 и Thpok и от белков TLE, что показывает: чрезмерно сильная репрессия Runx–TLE перекрывает обычную связь между специфичностью рецептора и выбором линейности.
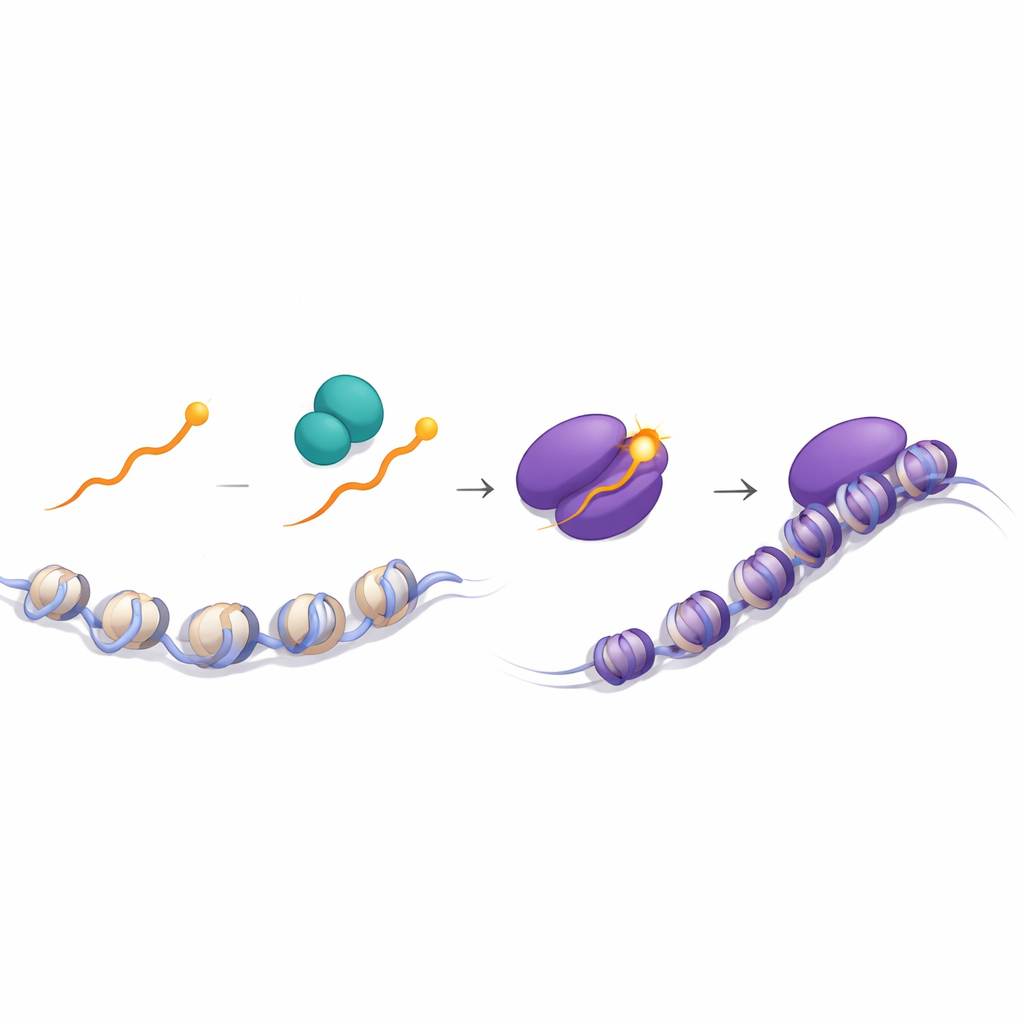
Фосфорилирующий переключатель, который улавливает внешние сигналы
Ключевой вывод был получен при изучении нормального хвоста Runx1. С помощью масс‑спектрометрии авторы обнаружили, что терминальный Y фосфорилируется — украшен фосфатной группой — значительно чаще в CD8‑тимоцитах, чем в CD4‑тимоцитах. Эта модификация сильно укрепляет взаимодействие Runx1 с TLE3. В клетках, лишённых тирозинкиназы Lck, эта фосфорилизация почти отсутствовала, что указывает на роль Lck и его партнёра Zap70, обоих ключевых передатчиков сигналов T‑клеточного рецептора. Имиджинговые эксперименты показали, что Runx1 встречается с Lck и Zap70 главным образом в цитоплазме, и такие встречи чаще происходят в клетках, сигнализированных через молекулы класса I. Структурные моделирования поддержали идею, что фосфорилированный Y, либо его замена на F или W, стабилизирует связывание Runx с WD40‑доменом TLE, способствуя сборке мощного репрессорного комплекса на сайленсерах.
От крошечной химической метки до иммунной идентичности
В совокупности исследование предлагает, что фосфорилирование одного терминального тирозина на белках Runx действует как чувствительный регулятор: в тимоцитах, сигнализированных через класс I, более высокая фосфорилизация эффективнее привлекает TLE к сайленсерам в генах Cd4 и Thpok, выключая их и фиксируя цитотоксическую CD8‑судьбу. В клетках, сигнализированных через класс II, низкая фосфорилизация делает связь Runx с TLE слабее, эти сайленсеры остаются неэффективными, и развивается программа хелперных CD4. Для неспециалиста смысл в том, что иммунная система может использовать крошечную химическую правку на одном конце белка, чтобы преобразовать тонкие рецепторные сигналы в безальтернативный выбор между двумя центральными типами T‑клеток, что иллюстрирует, насколько тонко настроенные молекулярные переключатели лежат в основе устойчивых решений в развитии.
Цитирование: Ogawa, C., Okuyama, K., Kojo, S. et al. Phosphorylation of Runx protein controls helper CD4+ T cell versus cytotoxic CD8+ T cell lineage choice. Nat Immunol 27, 799–811 (2026). https://doi.org/10.1038/s41590-026-02441-6
Ключевые слова: дифференцировка T‑клеток, фосфорилирование Runx, линейность CD4 против CD8, подавление генов, тимическое развитие