Clear Sky Science · zh
通过SLC7A4的pH响应性氨基酸运输的结构基础。
细胞为何关心酸性与氨基酸
我们的细胞持续摄入氨基酸——蛋白质的构件——以支持生长、大脑功能和免疫防御。本研究揭示了一种鲜为人知的人类蛋白SLC7A4如何将氨基酸亮氨酸搬运入细胞,以及这一过程在周围环境变为弱酸性时(常见于活跃或受压组织)如何增强。理解这种行为可加深我们对细胞如何感知营养的认识,并可能最终为针对依赖异常氨基酸利用的癌症和其他疾病提供新策略。
为亮氨酸隐藏的闸门
研究人员首先探问SLC7A4到底做什么。尽管它属于大型氨基酸转运蛋白家族,早期工作未能确定其运载的氨基酸。通过对纯化的人SLC7A4进行敏感的稳定性检测,团队发现亮氨酸——一种强烈影响细胞生长通路的支链氨基酸——特别能稳定该蛋白。当他们在缺少主要背景转运体的人细胞中表达SLC7A4时,细胞摄取的放射性亮氨酸远多于对照细胞,清楚地表明SLC7A4在细胞表面发挥亮氨酸转运功能。
外酸内输
接着,团队测试了细胞外酸度如何影响转运。他们将外部pH从接近中性的血液典型值降低到活跃或病变组织常见的弱酸性范围。随着环境变得更酸,SLC7A4介导的亮氨酸摄取在活细胞和包含纯化蛋白的人工膜囊泡中均显著增加。这表明SLC7A4是一种由细胞外质子浓度调节的pH响应型转运体。蛋白中的单个氨基酸——一个被称为Glu125的谷氨酸——被证明对该行为至关重要:将其改为非酸性残基在很大程度上消除了pH敏感性,而基本的转运能力仍然保留。
来自植物近缘的教训 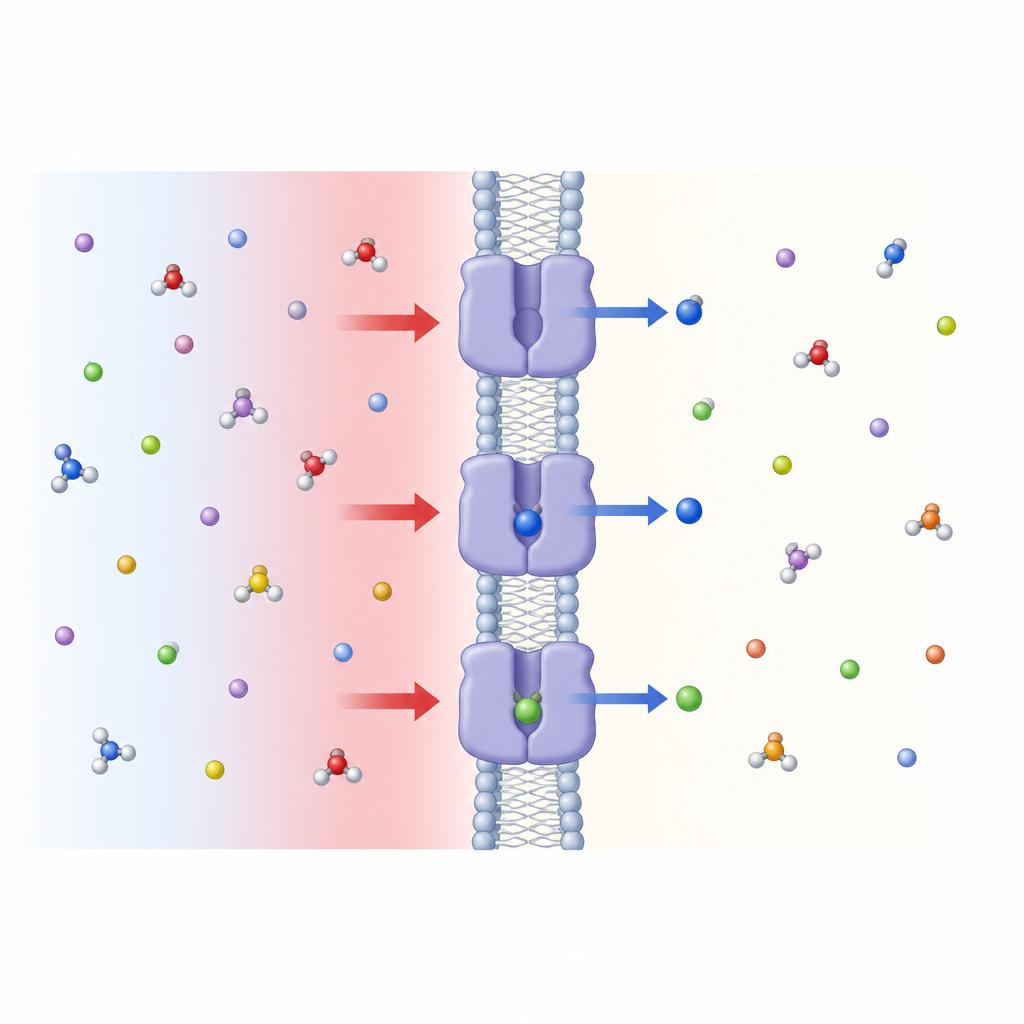
Figure 1. 酸性环境启动细胞膜上的一个闸门,使亮氨酸及相关氨基酸流入细胞。
为了在原子水平上观察这个闸门如何工作,研究人员转而研究来自拟南芥(Arabidopsis thaliana)的密切相关转运体AtCAT4,该蛋白更易于进行结构学研究。利用冷冻电子显微镜,他们捕获了AtCAT4在有无配体结合时的详细快照。该植物蛋白既能识别带正电的氨基酸也能识别亮氨酸,图像显示当配体位于结合口袋时,转运体的核心区域如何重排。计算机模拟表明,当氨基酸结合时,中央螺旋的一部分可以从平滑的螺旋变为弯曲形态,这种“诱导贴合”有助于将分子夹紧固定。这些运动与细菌近缘观察到的情形非常相似,指向该转运体家族的共同进化设计。
结合口袋如何在氨基酸中挑选亮氨酸 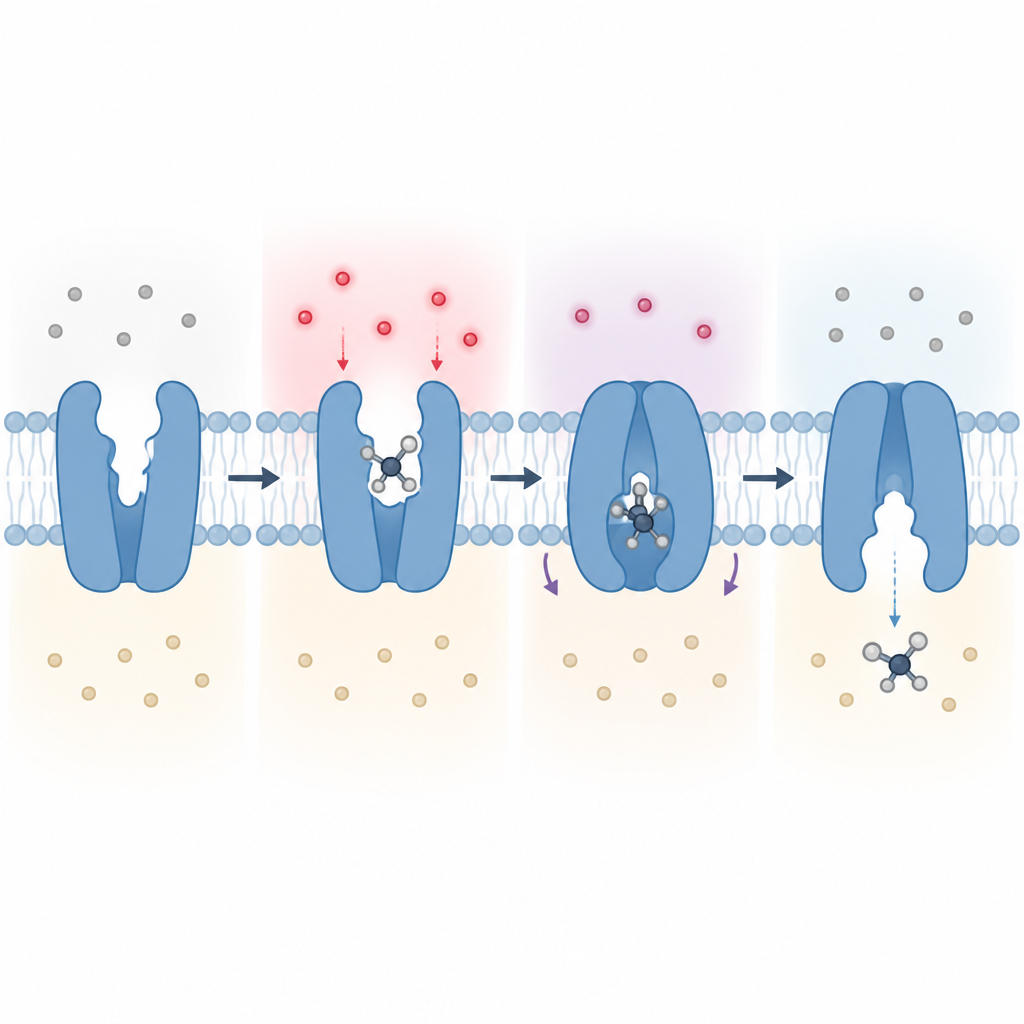
Figure 2. 一步步展示酸度如何调节蛋白的单个位点,从而重塑转运蛋白并将一分子亮氨酸穿过膜。
借助植物结构,团队构建了结合亮氨酸的人SLC7A4精细模型。在该模型中,亮氨酸的主链嵌入一个保守的口袋,而其支链侧链则塞入蛋白内部一簇紧密的疏水残基中。这个疏水口袋的微妙特征解释了为何SLC7A4偏好亮氨酸而非非常相似的氨基酸。通过将该口袋中的仅三个残基变为已知阳离子氨基酸载体的对应残基,研究人员能够翻转SLC7A4的偏好:突变体对精氨酸的结合与转运显著增强,而对亮氨酸的结合仅被适度削弱。这表明少数侧链就能作为开关,调节转运体偏好的氨基酸种类。
营养阀上的分子pH旋钮
结构学数据、模拟和细胞实验共同支持这样一个模型:Glu125位于pH敏感调控系统的核心。当细胞外为中性时,该残基大多不带电,帮助保持蛋白的向外开放构象。当环境变酸时,Glu125可被质子化,松动这种握持,使转运体更容易在向外和向内构象间循环,从而将亮氨酸运入细胞。因此这项工作将SLC7A4识别为质膜上的一个pH门控亮氨酸阀门,并以原子细节阐明其氨基酸偏好和酸敏性如何源自结合口袋中少数关键位置。
引用: Kolokouris, D., Bothra, A., Kato, T. et al. Structural basis for pH-responsive amino acid transport via SLC7A4.. Nat Commun 17, 4544 (2026). https://doi.org/10.1038/s41467-026-70956-5
关键词: 氨基酸转运, 亮氨酸, 膜蛋白, pH 感知, SLC7A4