Наши клетки постоянно импортируют аминокислоты — строительные блоки белков — чтобы поддерживать рост, работу мозга и иммунную защиту. В этом исследовании показано, как малоизвестный человеческий белок SLC7A4 помогает переносить аминокислоту лейцин в клетки и как этот процесс усиливается при небольшом подкислении окружающей среды, что часто происходит в активно работающих или стрессовых тканях. Понимание этого поведения может углубить наше представление о том, как клетки ощущают питательные вещества, и в перспективе помочь в разработке новых стратегий против рака и других заболеваний, зависящих от измененного использования аминокислот.
Скрытая «дверца» для лейцина
Исследователи начали с вопроса, чем именно занимается SLC7A4. Хотя он принадлежит к большой семье транспортёров аминокислот, предыдущие работы не смогли показать, какие именно аминокислоты он переносит. Используя чувствительный тест стабильности для очищенного человеческого SLC7A4, команда обнаружила, что лейцин — разветвлённая аминокислота, сильно влияющая на пути роста клеток — особенно эффективно стабилизирует белок. Когда SLC7A4 экспрессировали в человеческих клетках, лишённых одного из основных фоновых переносчиков, клетки захватывали значительно больше радиоактивного лейцина по сравнению с контролем, что явно указывает на то, что SLC7A4 функционирует как транспортер лейцина на поверхности клетки.
Кислота снаружи — движение внутри
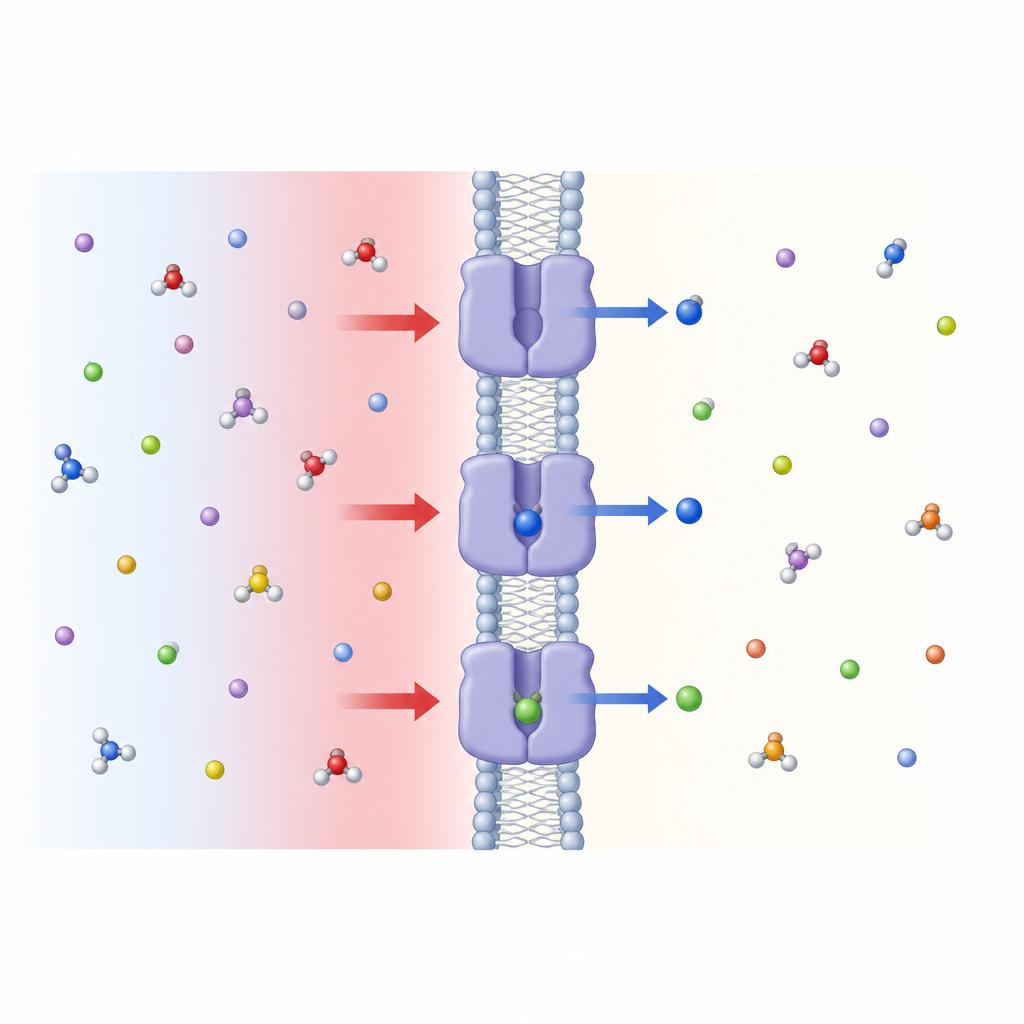
Далее команда проверила, как кислотность снаружи клетки влияет на транспорт. Они понизили внешний pH от почти нейтральных значений, типичных для крови, в сторону слабо кислого диапазона, часто встречающегося в активных или поражённых тканях. Захват лейцина через SLC7A4 резко вырос по мере подкисления среды, как в живых клетках, так и в искусственных мембранных пузырьках с очищенным белком. Это показало, что SLC7A4 — pH-реагирующий транспортер, чья активность регулируется концентрацией протонов снаружи клетки. Одна аминокислота в белке, глутамат с позицией 125 (Glu125), оказалась критичной для этого поведения: её замена на некислую остаток в значительной степени устраняла pH-чувствительность, при этом базовая транспортная способность оставалась.
Уроки от растительного родственника Figure 1. Кислая внешняя среда включает клеточную «дверцу» в мембране, через которую лейцин и родственные аминокислоты попадают в клетку.
Чтобы понять работу этой «дверцы» на атомном уровне, исследователи обратились к близкому транспортеру из растения Arabidopsis thaliana, называемому AtCAT4, который легче изучать структурно. С помощью крио-электронной микроскопии они получили детальные снимки AtCAT4 с лигандом и без него. Растительный белок распознаёт как положительно заряженные аминокислоты, так и лейцин, и изображения показали, как центральная область транспортёра перестраивается при наличии лиганда в участке связывания. Компьютерные симуляции показали, что часть центральной спирали может переключаться от гладкой конфигурации к изломанной при связывании аминокислоты — «индуцированное соответствие», которое помогает зажать молекулу на месте. Эти движения тесно напоминают наблюдаемые у бактериальных родственников, что указывает на общее эволюционное устройство этой семьи транспортеров.
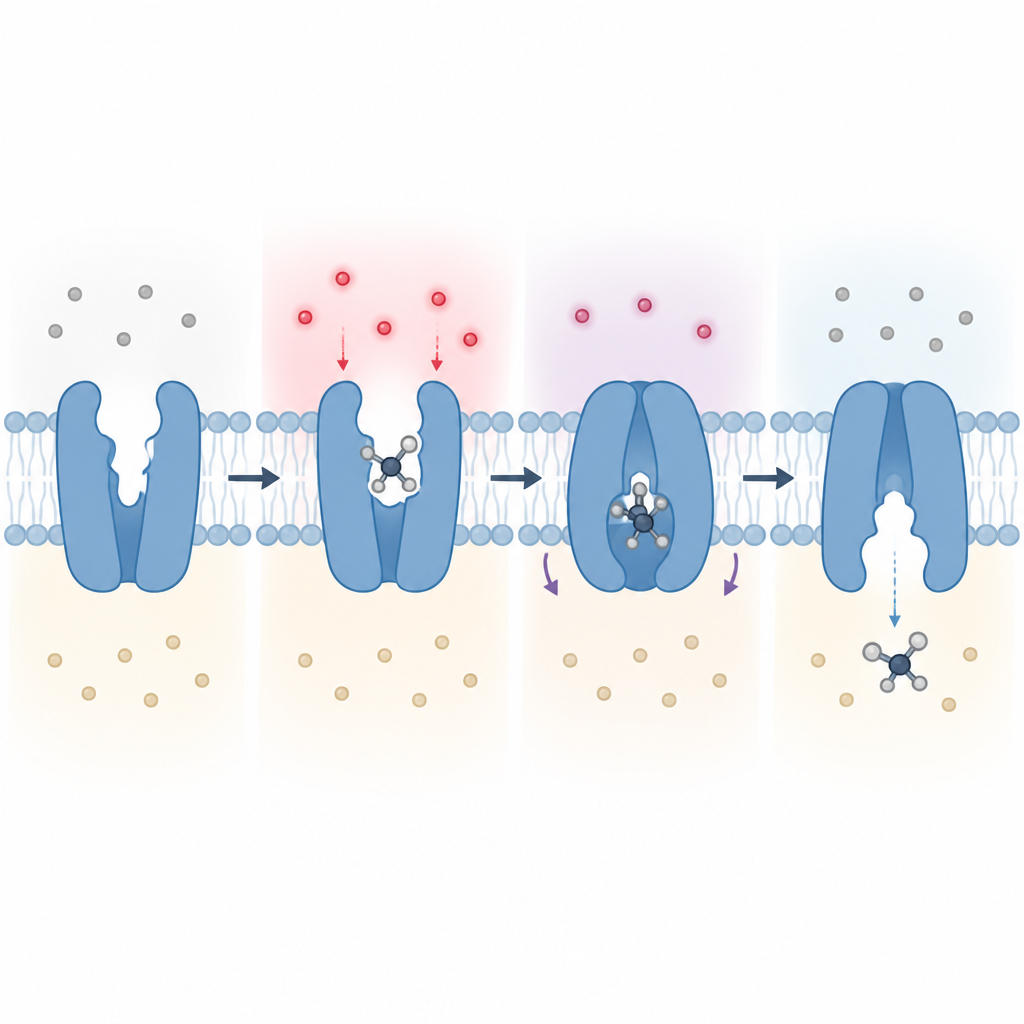
Как карман отбирает лейцин среди других аминокислот Figure 2. Пошаговое представление о том, как кислотность изменяет одну сайт-белок, перестраивая транспортер и перенося одну молекулу лейцина через мембрану.
Опираясь на растительную структуру, команда построила подробную модель человеческого SLC7A4 с связанным лейцином. В этой модели скелетная часть лейцина размещается в консервативном кармане, в то время как его разветвлённая боковая цепь заходит в плотное скопление гидрофобных остатков глубоко внутри белка. Тонкие особенности этого гидрофобного кармана объясняют, почему SLC7A4 предпочитает лейцин очень похожим аминокислотам. Изменив всего три остатка в этом кармане так, чтобы они соответствовали известному носителю положительно заряженных аминокислот, исследователи смогли изменить предпочтение SLC7A4: мутант сильнее связывал и транспортировал аргинин, при этом лишь умеренно ослабляя связывание лейцина. Это демонстрирует, что небольшой набор боковых цепей действует как переключатель, настраивающий, какие аминокислоты предпочитает транспортер.
Молекулярная «ручка pH» на клапане питательных веществ
Структурные данные, симуляции и клеточные эксперименты вместе поддерживают модель, в которой Glu125 находится в центре pH-чувствительной системы управления. Когда внешняя среда нейтральна, этот остаток в основном незаряжен и помогает удерживать части белка в открытой, ориентированной наружу конформации. При подкислении Glu125 может захватить протон, ослабляя это удержание и позволяя транспортеру легче циклировать между наружной и внутренней формами, перенося лейцин в клетку. Таким образом, работа идентифицирует SLC7A4 как pH-закрывающийся клапан для лейцина на плазматической мембране и подробно описывает на атомном уровне, как предпочтения по аминокислотам и чувствительность к кислотности возникают из нескольких ключевых позиций в его участке связывания.
Цитирование: Kolokouris, D., Bothra, A., Kato, T. et al. Structural basis for pH-responsive amino acid transport via SLC7A4..
Nat Commun17, 4544 (2026). https://doi.org/10.1038/s41467-026-70956-5
Ключевые слова: транспорт аминокислот, лейцин, мембранный белок, pH-сенсинг, SLC7A4