התאים שלנו מייבאים באופן קבוע חומצות אמינו, אבני הבניין של חלבונים, כדי להניע גדילה, פעילות מוחית והגנה חיסונית. המחקר הזה חושף כיצד חלבון אנושי מעט ידוע, SLC7A4, מסייע בהעברת חומצת האמינו לוצין אל התאים, וכיצד תהליך זה נעשה פעיל יותר כאשר הסביבה נהיית מעט חומצית, כפי שקורה לעתים קרובות ברקמות פעילות או עומסות. הבנת ההתנהגות הזו עשויה להעמיק את ידיעתנו על האופן שבו תאים חשים נוטריינטים ועלולה בסופו של דבר להנחות אסטרטגיות חדשות כנגד סרטן ומחלות אחרות התלויות בשינויי שימוש בחומצות אמינו.
שער חבוי ללוצין
החוקרים התחילו בשאלה מה SLC7A4 עושה בפועל. למרות ששייך למשפחה רחבה של מובילי חומצות אמינו, עבודות קודמות לא הצליחו להראות אילו חומצות אמינו הוא מעביר. באמצעות מבחן יציבות רגיש על SLC7A4 טהור אנושי, הצוות מצא שלוצין, חומצת אמינו מסועפת המשפיעה בחוזקה על מסלולי גדילה תאיים, הייתה יעילה במיוחד ביציבות החלבון. כאשר הביעו SLC7A4 בתאים אנושיים חסרי מוביל רקע מרכזי, התאים ייבאו הרבה יותר לוצין רדיואקטיבי מתאי בקרה, מה שמראה באופן ברור ש-SLC7A4 פועל כמוביל לוצין על פני שטח התא.
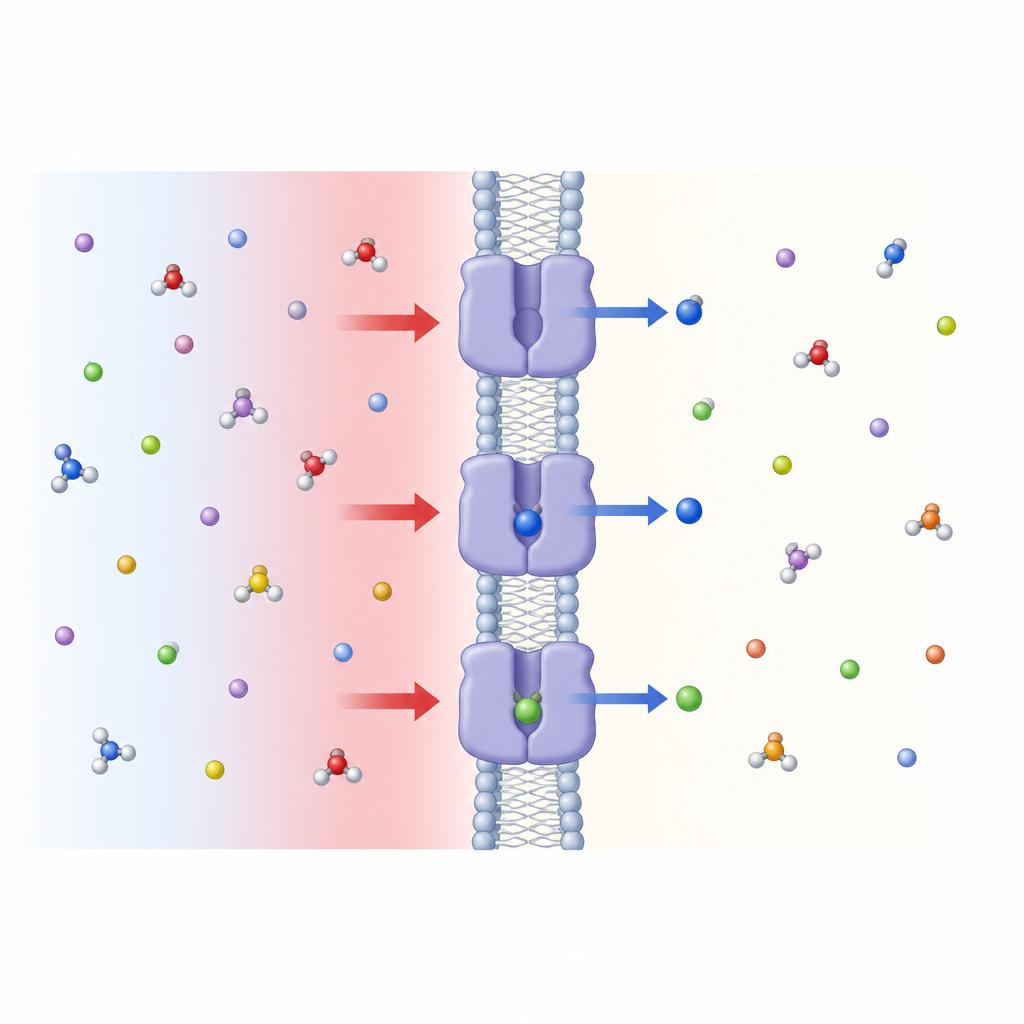
חומציות בחוץ, תנועה בפנים
לאחר מכן הקבוצה בדקה כיצד חומציות מחוץ התא משפיעה על הובלה. הם הורידו את ה-pH החיצוני מערכי ניטרל קרוב, אופייניים לדם, לטווח החומצי הקל הנפוץ ברקמות פעילות או חולות. ספיגת הלוצין דרך SLC7A4 עלתה בהתמדה ככל שהסביבה נעשתה חומצית יותר, הן בתאים חיים והן בבועות ממברנה מלאכותיות שהכילו את החלבון המטוהר. ממצא זה מראה כי SLC7A4 הוא מוביל רגיש ל-pH שהפעילות שלו מכויילת על פי ריכוז הפרוטונים מחוץ לתא. חומצת אמינו אחת בחלבון, גלוטמט בשם Glu125, התבררה כחשובה להתנהגות זו: שינוי שלה לשארית שאינה חומצית הסיר במידה רבה את הרגישות ל-pH תוך שמירה על יכולת ההובלה הבסיסית.
מסקנות מבקר קרוב של הצמח Figure 1. סביבה חומצית מפעילה שער בממברנת התא שמאפשר לזרום לתוך התא לוצין וחומצות אמינו קרובות.
כדי לראות כיצד השער עובד ברמת האטום, החוקרים פנו למוביל קרוב מצמח Arabidopsis thaliana, בשם AtCAT4, שקל יותר ללמוד אותו מבחינה מבנית. באמצעות מיקרוסקופ אלקטרונים בקירור הם תפסו תמונות מפורטות של AtCAT4 עם ובלי חומצת אמינו קשורה. חלבון הצמח מזהה גם חומצות אמינו בעלות מטען חיובי וגם לוצין, והתמונות הראו איך אזור ליבה במוביל מחדש מסודר כאשר ליגנד יושב בכיס הקשירה. סימולציות ממוחשבות הצביעו על כך שחלק מהאלפא-הליקס המרכזי יכול לעבור מצורת סליל חלקה לצורה מעוקמת כאשר חומצת אמינו נקשרת, "התאמה מושרת" שעוזרת לאחוז את המולקולה במקומה. התנועות האלה דומות מאוד לאלו הנצפות בקרובי משפחה חיידקיים, ומהוות עדות לעיצוב אבולוציוני משותף למשפחה זו של מובילים.
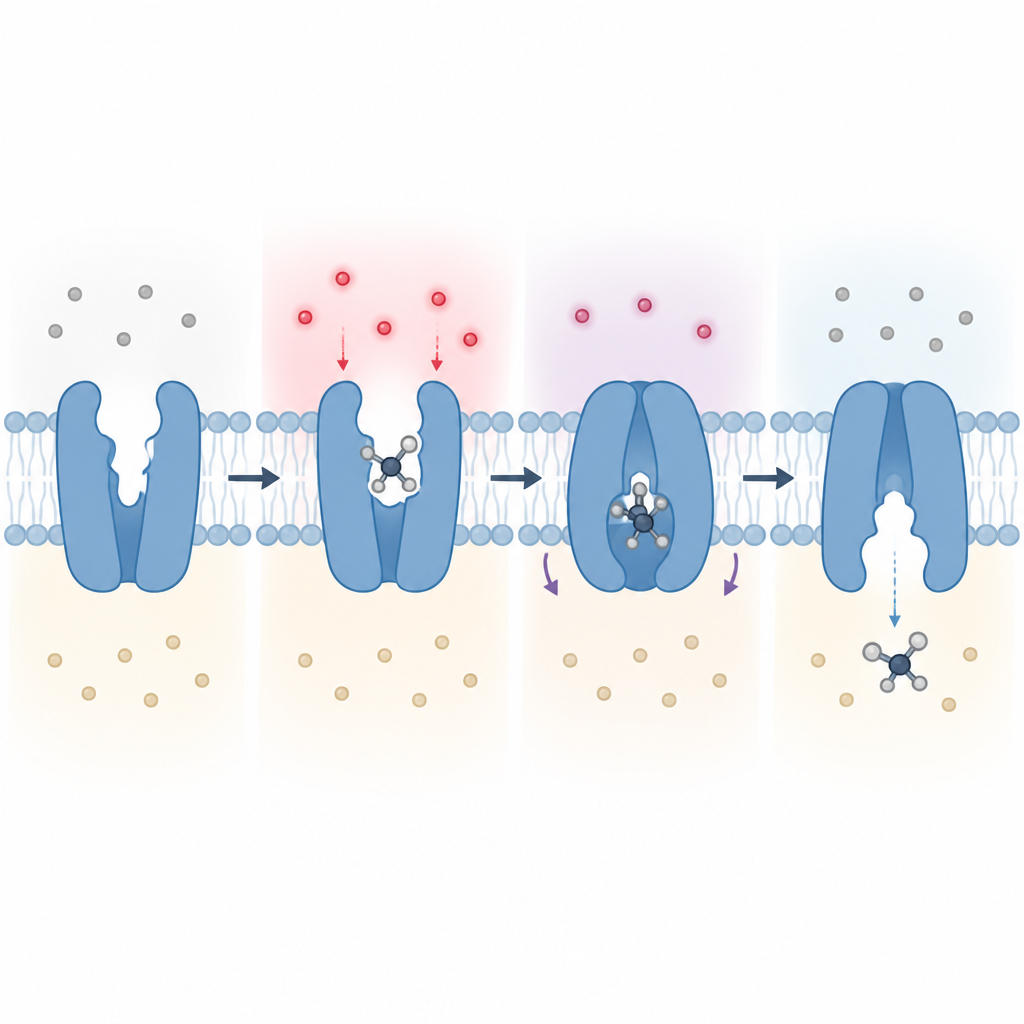
כיצד הכיס בוחר לוצין על פני חומצות אמינו אחרות Figure 2. תצפית שלב-אחר-שלב כיצד חומציות שמשתנה משפיעה על אתר יחיד בחלבון ומשנה את צורת המוביל כדי להעביר מולקולת לוצין אחת דרך הממברנה.
מצוידים במבנה הצמח, הקבוצה בנתה מודל מפורט של SLC7A4 האנושי עם לוצין קשור. במודל זה, השלד של הלוצין ננעץ בכיס משומר, בעוד ששרשרת הצד המסועפת שלו מתקפלת לתוך אשכול הדוק של שיירים הידרופוביים עמוק בחלבון. תכונות עדינות של כיס זה ההידרופובי מסבירות מדוע SLC7A4 מעדיף לוצין על פני חומצות אמינו דומות מאוד. על ידי שינוי שלוש שיירים בלבד בתוך הכיס כדי להתאים לאלה של נשא ידוע לחומצות אמינו טעונות חיובית, החוקרים הצליחו להפוך את ההעדפה של SLC7A4: המוטנט קישר והעביר ארגינין בעוצמה רבה יותר, בעוד שלוצין נחלש רק במידה מתונה. זה מראה שקבוצה קטנה של שיירי צד פועלת כמפסק שמכויל אילו חומצות אמינו המוביל מעדיף.
כפתור pH מולקולרי על שסתום ההזנה
הנתונים המבניים, הסימולציות וניסויי התאים יחד תומכים במודל שבו Glu125 ממוקם בלב מערכת בקרה רגישת pH. כאשר החוץ של התא ניטרלי, שיירה זו ברוב המקרים לא טעונה ועוזרת להחזיק חלקים מהחלבון בצורה פתוחה, הפונה החוצה. כאשר הסביבה נהיית חומצית, Glu125 יכולה לקלוט פרוטון, לשחרר את האחיזה ולאפשר למוביל להסתובב ביתר קלות בין מצבים הפונים החוצה והפונים פנימה כאשר הוא מעביר לוצין לתא. עבודה זו מזהה לכן את SLC7A4 כשסתום לוצין המופעל על ידי pH בממברנת הפלזמה ומפרטת ברמת האטום כיצד העדפות חומצות האמינו ורגישות החומציות שלו נובעות ממספר מוקדי מפתח בכיס הקשירה שלו.
ציטוט: Kolokouris, D., Bothra, A., Kato, T. et al. Structural basis for pH-responsive amino acid transport via SLC7A4..
Nat Commun17, 4544 (2026). https://doi.org/10.1038/s41467-026-70956-5