Clear Sky Science · zh
UHRF1 的选择性细胞定位保障哺乳动物合子基因组激活与早期胚胎发育
把生命最早的步骤保持在正轨上
每一个哺乳动物的生命都始于受精卵,该受精卵必须迅速学会运行自身的遗传程序。一个关键的早期里程碑是胚胎首次启动自身基因的时刻,称为合子基因组激活。该研究提出了一个看似简单的问题:胚胎如何在这一关键时刻阻止某些强效的 DNA 结合蛋白干扰?通过追踪这些蛋白在小鼠胚胎中的分布,研究者揭示了一套“门控”机制,将它们在正确的时间保持在合适的细胞区室,从而有助于确保正常发育。
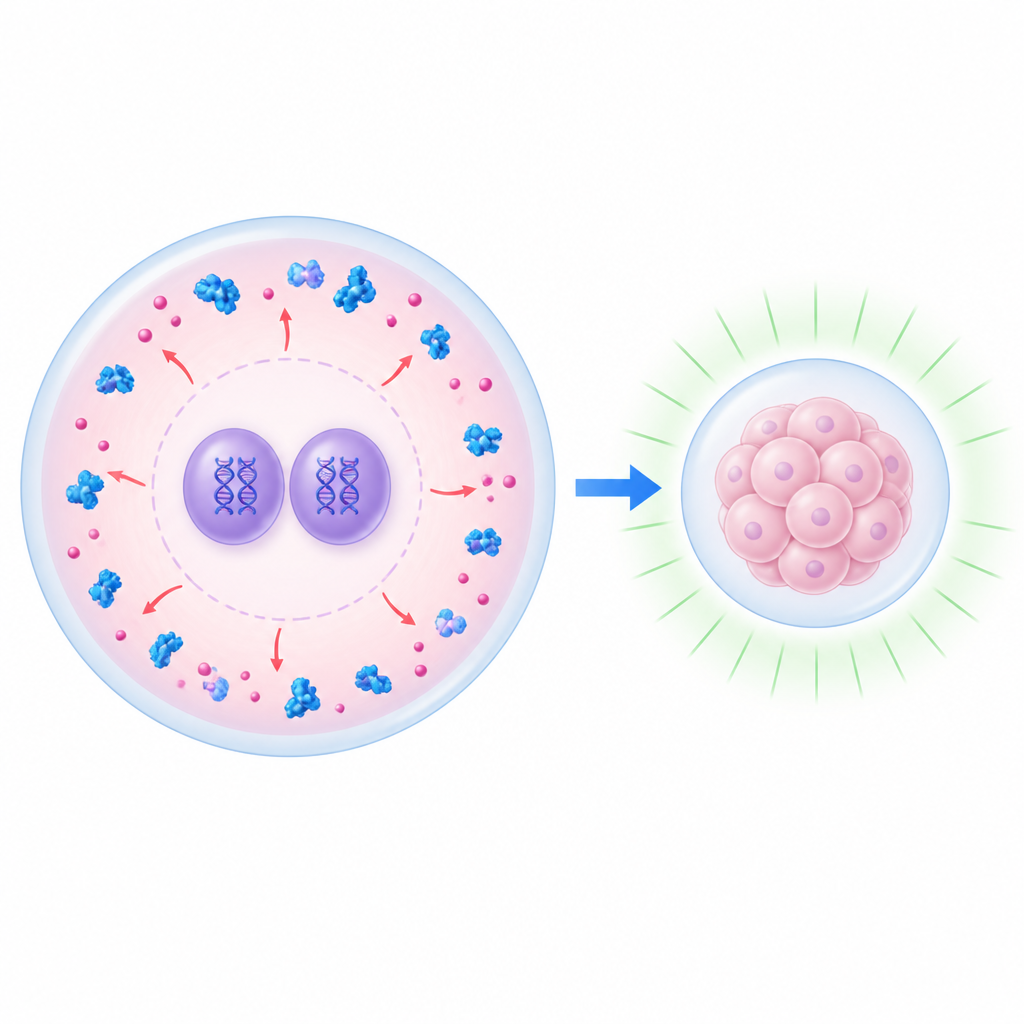
早期发育核心处的门控
在受精后的最初几小时内,胚胎仍依赖母体留下的分子。其中包括两种蛋白 UHRF1 和 DNMT1,长期以来被认为有助于维持通常使基因沉默的 DNA 化学标记。令人意外的是,在健康的小鼠胚胎中,这些蛋白在很大程度上被排除在新形成的细胞核之外——父母的基因组就位于这些核内。研究团队使用缺失一种母体因子 NLRP14 的小鼠作为模型,NLRP14 通常有助于在受精后将 UHRF1 与 DNMT1 保留在细胞质中。缺少 NLRP14 时,这两种蛋白涌入细胞核,合子基因组激活被强烈阻断,胚胎停滞在二细胞期,这表明这些蛋白在细胞内的位置会决定早期发育的成败。
错位蛋白如何锁定基因组
为了解核内 UHRF1 的实际作用,研究者绘制了其在基因组上的结合位点并测量 DNA 的包装紧密程度。当 UHRF1 在 Nlrp14 缺陷胚胎的细胞核中积累时,它强烈结合许多重复 DNA 序列,包括称为 LINE1 的长散在重复元件和某些长末端重复(LTR)片段。这些位点的可及性降低,仿佛在染色质上加了额外的锁。同时,许多应当被激活的早期胚胎基因仍然保持沉默。研究显示,这种有害结合部分依赖于 DNA 甲基化——UHRF1 能识别这种化学标记,提示蛋白和这些标记过量共存会在基因组本应打开时将其固定在受抑状态。
将因果关系区分开来
由于 NLRP14 可能影响多种分子,作者构建了双突变小鼠以精确定位 UHRF1 的特定作用。与此同时删除 UHRF1 和 NLRP14 使许多胚胎突破二细胞阻滞并恢复大多数早期基因的活性,即便受精后通常会被清除的大量 DNA 甲基化仍然存在。相比之下,同时删除 DNMT1 与 NLRP14,或化学性阻断 UHRF1 识别甲基化 DNA 的能力,虽然能促进染色质开放并恢复相当比例的早期基因,但并未完全挽救发育。这些比较表明,过量的核内 UHRF1,而非单纯的全局 DNA 甲基化,是抑制胚胎首轮基因激活的主要刹车。
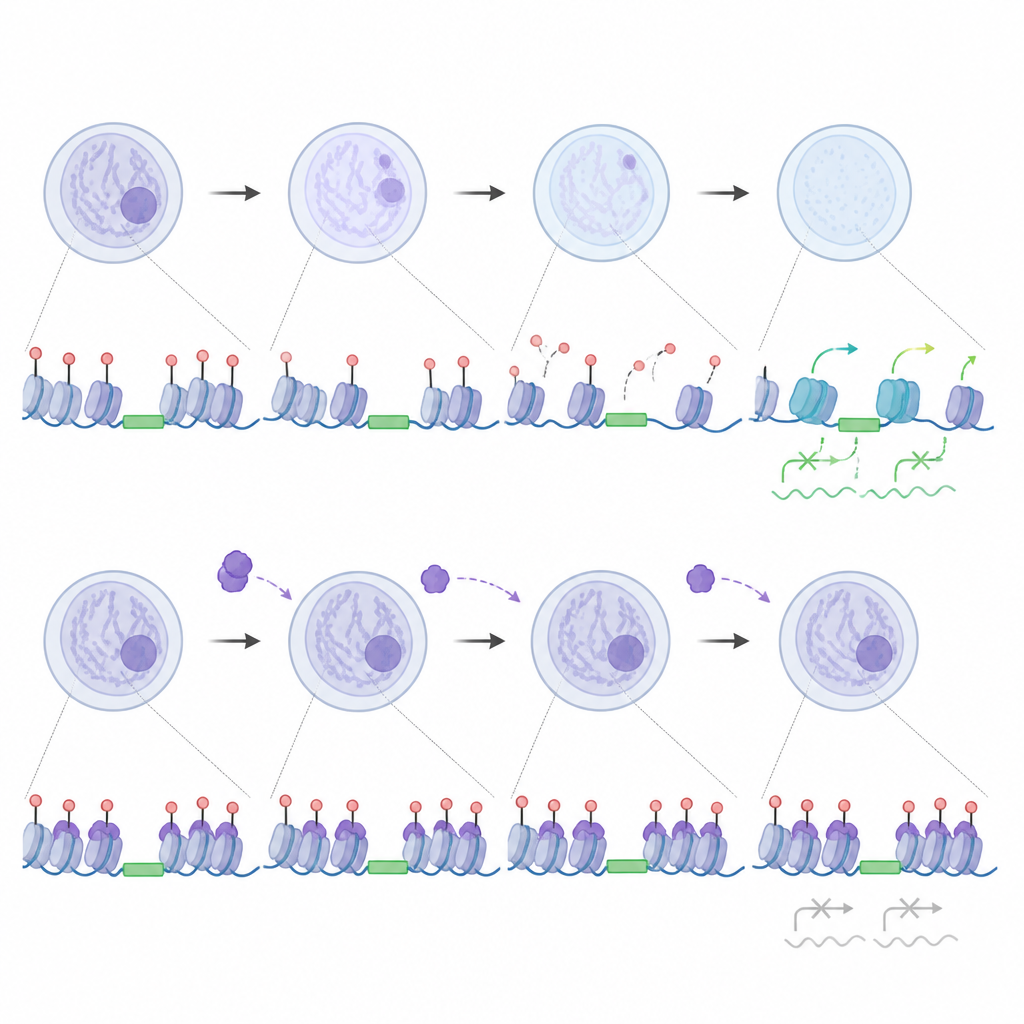
调节“跳跃基因”而不是一律沉默它们
这项工作也改变了我们对所谓跳跃基因的看法。某些可移动的 DNA 元件,尤其是某些 LINE1 家族,当它们被转录时,实际上有助于触发合子基因组激活。研究者发现,当 UHRF1 和 DNMT1 被排除在细胞核之外时,这些 LINE1 区域失去甲基化,免受大量 UHRF1 结合,变得具有活性,从而反过来支持更开放的染色质和适当的基因切换。与此同时,少量正常进入细胞核的 UHRF1 会结合特定的 LTR 亚型,使其保持甲基化并保持沉默。在缺失 UHRF1 的胚胎中,这些特定重复序列异常激活,并与染色质开放性的细微变化相关,表明胚胎通常使用 UHRF1 对一小部分元件施加精细的制动,同时允许其他元件助力发育。
为何这种细胞门控很重要
对非专业读者而言,主要信息是早期胚胎必须谨慎控制不仅是产生哪些蛋白,还要精确控制这些蛋白在细胞内的去向。该研究表明,在受精后立即将 UHRF1 与 DNMT1 排除在细胞核之外,可以防止它们过度收紧基因组并关闭有益的 DNA 重复序列。与此同时,一小部分位置得当的 UHRF1 帮助让少数顽固的重复保持沉默。总体而言,这些基于位置的规则让胚胎在保护基因组与唤醒自身基因之间取得平衡。鉴于 UHRF1 及相关机制在许多动物中是保守的,理解这种空间控制可能有助于阐明生育力、早期发育以及表观遗传信息在生命开端如何被重置的普遍原理。
引用: Yan, R., Cheng, X., Long, X. et al. Selective cellular localization of UHRF1 safeguards mammalian zygotic genome activation and early embryonic development. Cell Discov 12, 38 (2026). https://doi.org/10.1038/s41421-026-00896-3
关键词: 合子基因组激活, UHRF1, DNA 甲基化, LINE1 元件, 早期胚胎发育