Clear Sky Science · it
Localizzazione cellulare selettiva di UHRF1 tutela l’attivazione genica zigotica e lo sviluppo embrionale precoce nei mammiferi
Mantenere i primissimi passi della vita sul giusto percorso
Ogni vita mammifera inizia con un uovo fecondato che deve in fretta imparare a eseguire il proprio programma genetico. Una tappa cruciale precoce è quando l’embrione accende per la prima volta i propri geni, un processo chiamato attivazione del genoma zigotico. Questo studio pone una domanda apparentemente semplice: come fa l’embrione a impedire che certe potenti proteine leganti il DNA intralcino questo momento critico? Tracciando queste proteine negli embrioni di topo, i ricercatori scoprono un sistema di controllo che le mantiene nel compartimento cellulare giusto al momento giusto, contribuendo a garantire uno sviluppo normale.
Un cancello al centro dello sviluppo precoce
Nelle primissime ore dopo la fecondazione, l’embrione dipende ancora da molecole lasciate dalla madre. Tra queste vi sono due proteine, UHRF1 e DNMT1, note da tempo per il loro ruolo nel mantenere i marchi chimici sul DNA che in genere mantengono i geni spenti. Sorprendentemente, negli embrioni di topo sani queste proteine sono in gran parte tenute fuori dai nuclei di nuova formazione, dove risiedono i genomi parentali. Il gruppo ha utilizzato topi privi di un fattore materno chiamato NLRP14, che normalmente aiuta a trattenere UHRF1 e DNMT1 nel citoplasma circostante dopo la fecondazione. Senza NLRP14, entrambe le proteine affluiscono nei nuclei, l’attivazione del genoma zigotico è fortemente bloccata e gli embrioni si arrestano allo stadio a due cellule, indicando che la posizione di queste proteine all’interno della cellula può compromettere lo sviluppo precoce.
Come proteine mal posizionate serrano il genoma
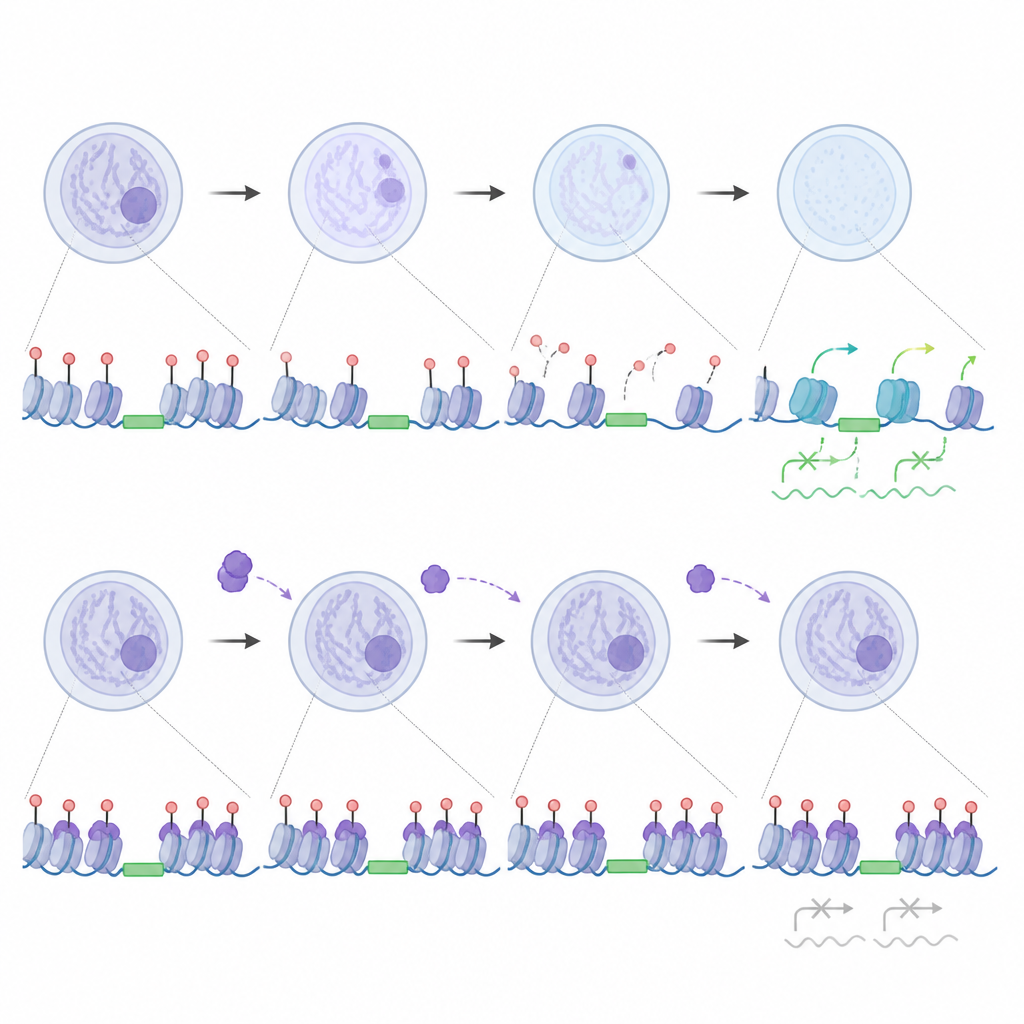
Per capire cosa faccia realmente UHRF1 quando è nel nucleo, i ricercatori hanno mappato i suoi siti di legame sul genoma e misurato quanto strettamente il DNA è impacchettato. Quando UHRF1 si accumula nei nuclei degli embrioni privi di Nlrp14, si lega intensamente a molte sequenze di DNA ripetute, inclusi elementi intercalati lunghi noti come LINE1 e alcuni segmenti a terminale lungo (LTR). Questi siti diventano meno accessibili, come se fossero stati aggiunti serrature extra alla cromatina. Allo stesso tempo, molti geni embrionali precoci che dovrebbero attivarsi restano silenti. Lo studio mostra che questo legame dannoso dipende in parte dalla metilazione del DNA, un marchio chimico riconosciuto da UHRF1, suggerendo che un’eccessiva presenza sia della proteina sia di questi marchi può congelare il genoma in uno stato reprimente proprio quando deve aprirsi.
Separare causa ed effetto
Poiché NLRP14 potrebbe influenzare molte molecole, gli autori hanno generato topi doppi‑mutanti per individuare il ruolo specifico di UHRF1. La rimozione di UHRF1 insieme a NLRP14 ha permesso a molti embrioni di superare il blocco a due cellule e ha ripristinato l’attività della maggior parte dei geni precoci, nonostante gran parte della metilazione del DNA che normalmente viene eliminata dopo la fecondazione fosse ancora presente. Al contrario, eliminare DNMT1 insieme a NLRP14, o bloccare chimicamente la capacità di UHRF1 di riconoscere il DNA metilato, ha facilitato l’apertura della cromatina e riportato all’attività una larga frazione di geni precoci ma non ha completamente recuperato lo sviluppo. Questi confronti rivelano che l’eccesso di UHRF1 nucleare, più che la sola metilazione globale del DNA, è il freno dominante sulla prima ondata di attivazione genica dell’embrione.
Regolare i geni saltellanti invece di zittirli tutti
Il lavoro cambia anche il modo in cui pensiamo ai cosiddetti geni saltellanti. Alcuni elementi di DNA mobili, in particolare certe famiglie di LINE1, aiutano in realtà a innescare l’attivazione del genoma zigotico quando vengono trascritti. I ricercatori hanno trovato che quando UHRF1 e DNMT1 sono esclusi dal nucleo, queste regioni LINE1 perdono metilazione, restano libere da un marcato legame di UHRF1 e si attivano, il che a sua volta favorisce una cromatina più aperta e il corretto switching genico. Allo stesso tempo, una piccola quantità di UHRF1 che normalmente entra nei nuclei si lega a sottotipi specifici di LTR mantenendo la loro metilazione e restando silenziosi. Negli embrioni privi di UHRF1, questi particolari ripetuti diventano anormalmente attivi e sono associati a cambiamenti sottili nell’apertura della cromatina, suggerendo che l’embrione usa normalmente UHRF1 come freno finemente tarato su un gruppo selezionato di elementi permettendo ad altri di supportare lo sviluppo.
Perché questa regolazione cellulare è importante
Per un non specialista, il messaggio principale è che gli embrioni precoci devono controllare con cura non solo quali proteine producono, ma esattamente dove queste proteine si localizzano nella cellula. Questo studio mostra che escludere UHRF1 e DNMT1 dal nucleo subito dopo la fecondazione impedisce loro di stringere eccessivamente il genoma e di spegnere ripetuti di DNA utili. Allo stesso tempo, una piccola frazione di UHRF1 ben posizionata aiuta a mantenere alcuni ripetuti ostinati silenziosi. Nel loro insieme, queste regole basate sulla posizione permettono all’embrione di bilanciare la protezione del genoma con la necessità di risvegliare i propri geni. Poiché UHRF1 e i meccanismi correlati sono conservati in molti animali, comprendere questo controllo spaziale può illuminare principi generali della fertilità, dello sviluppo precoce e di come l’informazione epigenetica venga resettata all’inizio della vita.
Citazione: Yan, R., Cheng, X., Long, X. et al. Selective cellular localization of UHRF1 safeguards mammalian zygotic genome activation and early embryonic development. Cell Discov 12, 38 (2026). https://doi.org/10.1038/s41421-026-00896-3
Parole chiave: attivazione del genoma zigotico, UHRF1, metilazione del DNA, elementi LINE1, sviluppo embrionale precoce