Clear Sky Science · pl
Selektywna lokalizacja komórkowa UHRF1 zabezpiecza aktywację genomu zygoty i wczesny rozwój zarodkowy ssaków
Utrzymanie najwcześniejszych etapów życia na właściwym torze
Każde życie ssaka zaczyna się od zapłodnionego jaja, które musi szybko nauczyć się samodzielnie uruchamiać swój program genetyczny. Kluczowym wczesnym etapem jest moment, gdy zarodek po raz pierwszy włącza własne geny — proces zwany aktywacją genomu zygoty. To badanie stawia pozornie proste pytanie: jak zarodek powstrzymuje pewne silne białka wiążące DNA przed przeszkadzaniem w tym krytycznym momencie? Śledząc te białka w embrionach myszy, badacze odkryli system bramkowania, który utrzymuje je w odpowiednim przedziale komórkowym we właściwym czasie, pomagając zapewnić prawidłowy rozwój.
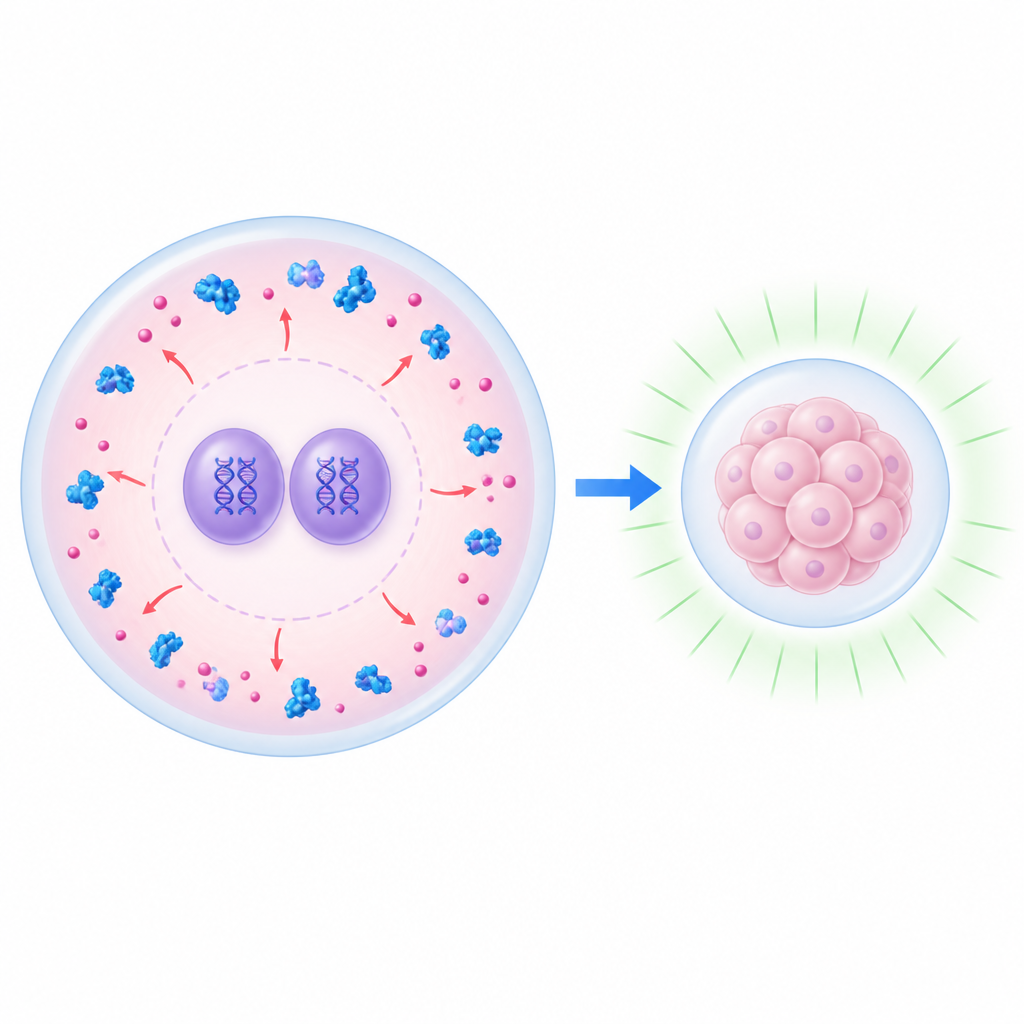
Bramka w sercu wczesnego rozwoju
W pierwszych godzinach po zapłodnieniu zarodek nadal polega na cząsteczkach pozostawionych przez matkę. Wśród nich są dwa białka, UHRF1 i DNMT1, od dawna znane z utrzymywania chemicznych znaczników na DNA, które zazwyczaj utrzymują geny w stanie nieaktywnym. Zaskakująco, u zdrowych embrionów myszy białka te w dużej mierze są utrzymywane poza świeżo utworzonymi jądrami, gdzie znajdują się genomy rodzicielskie. Zespół użył myszy pozbawionych czynnika macierzy zwanego NLRP14, który normalnie pomaga zatrzymywać UHRF1 i DNMT1 w otaczającej cytoplazmie po zapłodnieniu. Bez NLRP14 oba białka zalewają jądra, aktywacja genomu zygoty jest silnie zablokowana, a embriony zatrzymują się na etapie dwukomórkowym, co pokazuje, że miejsce występowania tych białek wewnątrz komórki może decydować o powodzeniu wczesnego rozwoju.
Jak niewłaściwie zlokalizowane białka blokują genom
Aby zobaczyć, co jądrowy UHRF1 faktycznie robi, badacze zmapowali miejsca jego wiązania w całym genomie i zmierzyli, jak ciasno upakowane jest DNA. Kiedy UHRF1 gromadzi się w jądrach embrionów pozbawionych Nlrp14, silnie wiąże się z wieloma sekwencjami powtórzonymi, w tym z długimi elementami przerywanymi znanymi jako LINE1 oraz z niektórymi odcinkami końcowymi typu LTR. Te miejsca stają się mniej dostępne, jakby do chromatyny dołożono dodatkowe zamki. Jednocześnie wiele wczesnych genów embrionalnych, które powinny się aktywować, pozostaje cichych. Badanie pokazuje, że to szkodliwe wiązanie zależy częściowo od metylacji DNA — chemicznego znaku, który UHRF1 potrafi rozpoznać — co sugeruje, że nadmiar zarówno białka, jak i tych znaków może zamrozić genom w stanie represyjnym dokładnie wtedy, gdy powinien się otwierać.
Oddzielanie przyczyny od skutku
Ponieważ NLRP14 mógł wpływać na wiele cząsteczek, autorzy stworzyli myszy podwójnie mutantne, by wyizolować specyficzną rolę UHRF1. Usunięcie UHRF1 wraz z NLRP14 pozwoliło wielu embrionom przejść blok dwukomórkowy i przywróciło aktywność większości wczesnych genów, nawet jeśli znaczna część metylacji DNA, która normalnie jest usuwana po zapłodnieniu, pozostała. W przeciwieństwie do tego, usunięcie DNMT1 razem z NLRP14, lub chemiczne zablokowanie zdolności UHRF1 do rozpoznawania metylowanego DNA, ułatwiło otwieranie chromatyny i przywróciło dużą część wczesnych genów, ale nie uratowało w pełni rozwoju. Te porównania pokazują, że nadmierna ilość jądrowego UHRF1, a nie sama globalna metylacja DNA, jest dominującą blokadą pierwszej fali aktywacji genów zarodka.
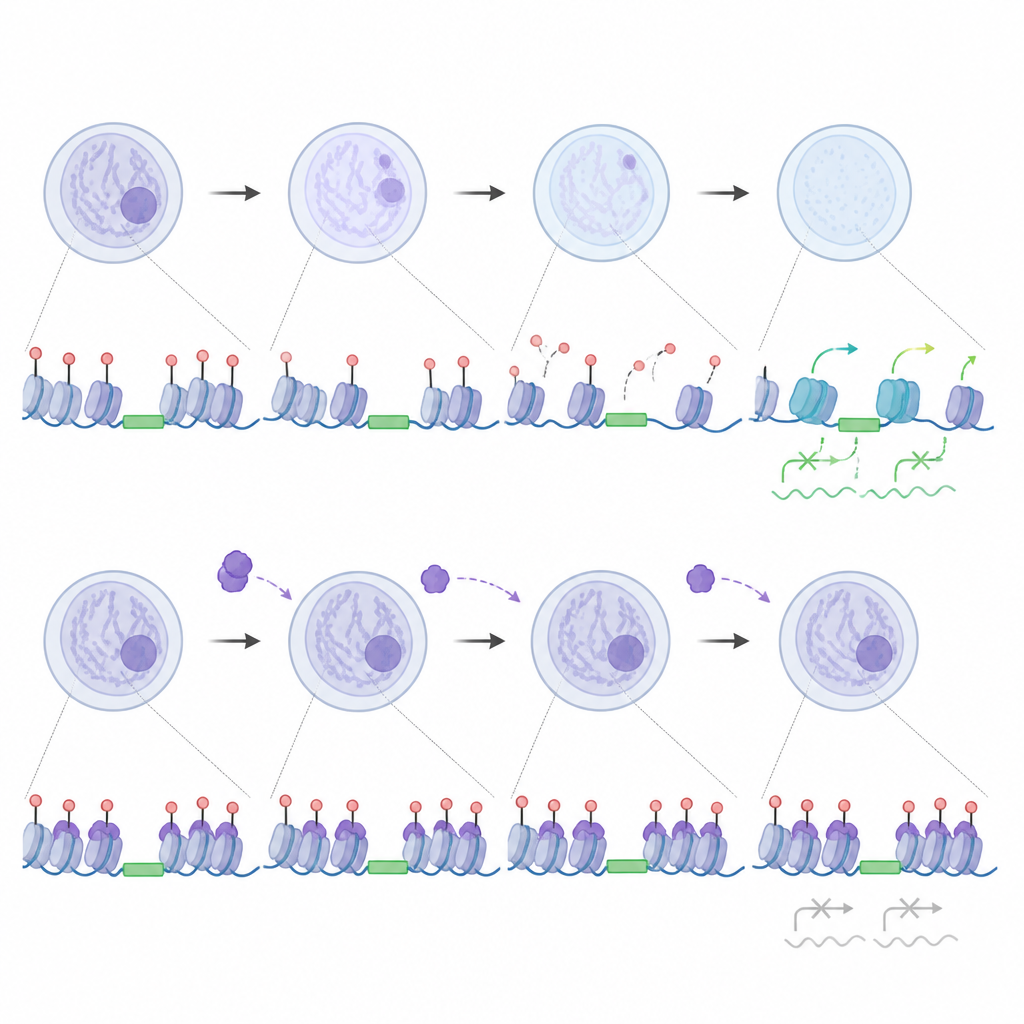
Strojenie „skaczących” genów zamiast ich powszechnego wyciszania
Praca zmienia też sposób, w jaki myślimy o tzw. genach skaczących. Niektóre ruchome elementy DNA, zwłaszcza niektóre rodziny LINE1, rzeczywiście pomagają uruchomić aktywację genomu zygoty, gdy są transkrybowane. Badacze odkryli, że gdy UHRF1 i DNMT1 są wykluczone z jądra, obszary LINE1 tracą metylację, pozostają wolne od silnego wiązania UHRF1 i stają się aktywne, co z kolei wspiera bardziej otwartą chromatynę i prawidłowe przełączanie genów. Jednocześnie niewielka ilość UHRF1, która normalnie wchodzi do jąder, wiąże specyficzne podtypy LTR, które zachowują metylację i pozostają ciche. W embrionach pozbawionych UHRF1 te konkretne powtórzenia stają się nieprawidłowo aktywne i łączą się z subtelnymi zmianami w dostępności chromatyny, co sugeruje, że zarodek normalnie używa UHRF1 jako precyzyjnego hamulca dla wybranej grupy elementów, jednocześnie pozwalając innym wspierać rozwój.
Dlaczego ta komórkowa kontrola lokalizacji ma znaczenie
Dla osoby niezwiązanej z dziedziną główne przesłanie jest takie, że wczesne embriony muszą starannie kontrolować nie tylko to, które białka produkują, ale także dokładnie, dokąd te białka trafiają w komórce. Badanie pokazuje, że wykluczenie UHRF1 i DNMT1 z jądra bezpośrednio po zapłodnieniu zapobiega nadmiernemu „dokręcaniu” genomu i wyłączaniu pomocnych powtórzeń DNA. Równocześnie niewielka, dobrze ulokowana frakcja UHRF1 pomaga utrzymać kilka uporczywych powtórzeń w spokoju. Razem te reguły oparte na lokalizacji pozwalają zarodkowi zrównoważyć ochronę genomu z potrzebą „obudzenia” własnych genów. Ponieważ UHRF1 i pokrewne mechanizmy są zachowane u wielu zwierząt, zrozumienie tej kontroli przestrzennej może przybliżyć ogólne zasady dotyczące płodności, wczesnego rozwoju i tego, jak informacja epigenetyczna jest resetowana na samym początku życia.
Cytowanie: Yan, R., Cheng, X., Long, X. et al. Selective cellular localization of UHRF1 safeguards mammalian zygotic genome activation and early embryonic development. Cell Discov 12, 38 (2026). https://doi.org/10.1038/s41421-026-00896-3
Słowa kluczowe: aktywacja genomu zygoty, UHRF1, metylacja DNA, elementy LINE1, wczesny rozwój zarodkowy