Clear Sky Science · tr
Kromatina özgü mekanizmalar yönelime özgü sınıf değiştirme rekombinasyonunu belirler
Vücudumuz Antikorları Nasıl İnce Ayarlar
Enfeksiyonlarla savaşırken bağışıklık sistemimiz tek tür bir antikora bağlı kalmaz. Akciğerlerdeki virüslerden bağırsaktaki bakterilere kadar farklı tehditlere daha iyi uyum sağlamak için antikor moleküllerinin parçalarını değiştirebilir. Bu makale, bağışıklık hücreleri içindeki DNA’nın fiziksel katlanmasının bu değiştirme sürecini nasıl sessizce yönlendirdiğini araştırıyor; böylece antikor değişikliklerinin etkili, doğru ve vücudun ihtiyaçlarına uygun olmasını sağlıyor.
Antikor Genleri İçindeki Anahtar Panel
Antikorlar, modüler parçalar gibi yeniden düzenlenip uyarlanabilen gen segmentlerinden yapılır. Bir B hücresinin yaşamının erken döneminde, bir dizi değişiklik antikorun mikrobu tanıyan “baş” bölümünü oluşturur. Daha sonra, sınıf değiştirme denilen ikinci bir süreç “kuyruk” bölgesini değiştirir; bu, antikorun vücutta nasıl davrandığını—kan dolaşımında mı kalacağı, bağırsağı mı kaplayacağı, yoksa mukozal yüzeyleri mi geçeceği—belirler. Bu değişim antikor ağır zincir geninde olur; burada varsayılan bir segment, birkaç alternatiften biriyle değiştirilir. DNA’yı kesen özel bir enzim bu değişime izin verdiği uzun zamandır bilinse de, bu kesimlerin neden genellikle tek bir yönelimde yeniden birleştikleri—dolayısıyla kullanılabilir bir antikor üretilip kırık veya sessiz bir ürün yerine işlevsel sonuçlar verdiği—halen merak konusudur.
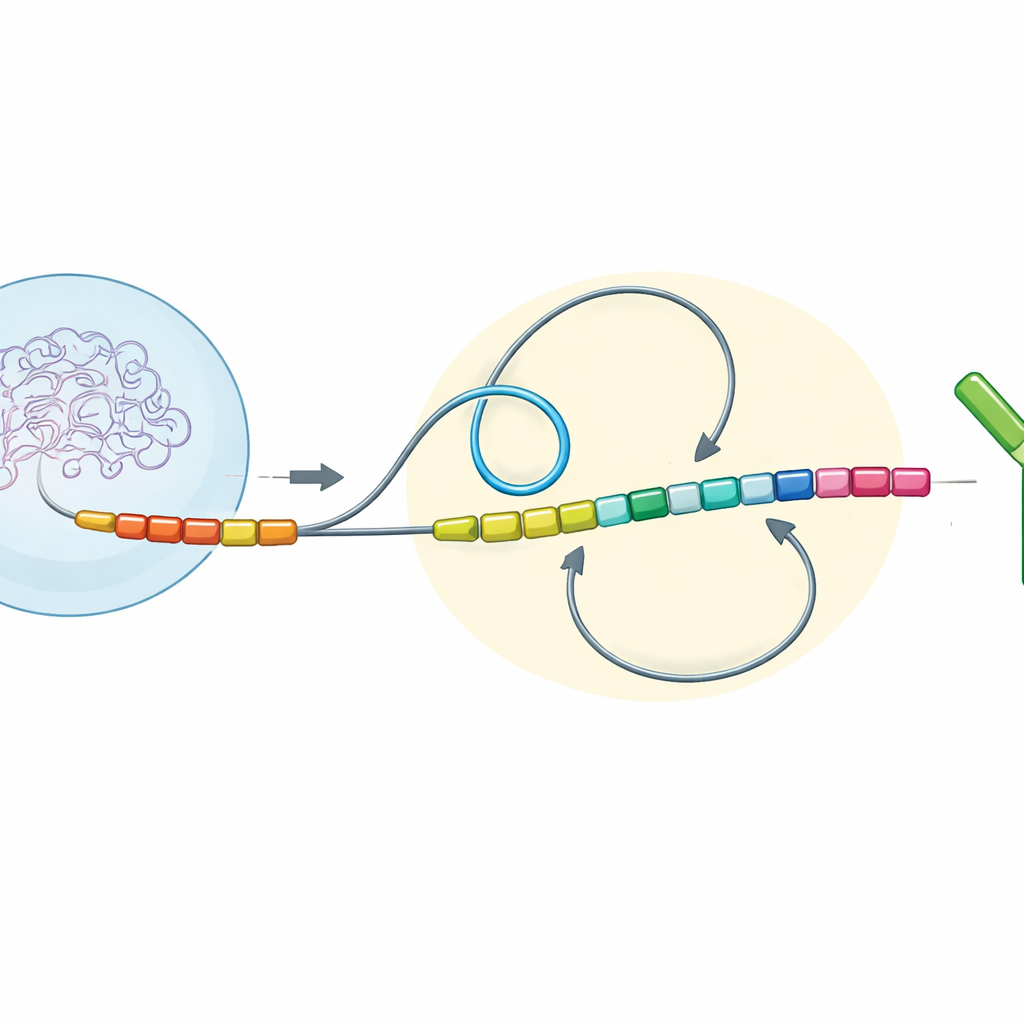
DNA Halkaları ve Yön Sonucu Şekillendirir
Yazarlar birçok omurgalı türü boyunca antikor gen bölgelerini incelediler ve ardından bu bölgelerin değiştirilmiş versiyonlarını laboratuvarda büyütülen fare bağışıklık hücrelerinde yeniden inşa ettiler. DNA’nın üç basit ama güçlü özelliğine odaklandılar: farklı segmentlerin okunduğu yön (transkripsiyon yönü), kromozom boyunca ne kadar uzakta oldukları (kromatin mesafesi) ve aynı üç boyutlu mahalle (kromatin bölgesi) içinde mi yoksa ayrı bölümlerde mi yer aldıkları. Birlikte, bu kombinasyona “anahtar topolojik konfigürasyonu” adını veriyorlar. Belirli sabit bölge segmentlerini kesin olarak çevirerek, taşıyarak veya birleştirerek bu değişikliklerin sınıf değiştirme sırasında kırık DNA uçlarının nasıl yeniden birleştirildiğini nasıl etkilediğini izleyebildiler.
Uzun Halkaların Egemen Olduğu Durumlar vs. Yerel Çarpışmaların Kazandığı Zamanlar
Fareler ve insanlar gibi memelilerde, çoğu antikor “kuyruk” segmenti başlangıç segmentiyle aynı yönde dizilmiş ve gen üzerinde göreli olarak uzak aralıklarla yerleşmiştir. Bu düzenekte, bir protein kompleksi DNA’yı büyük halkalar halinde sarar ve uzak parçaları tercih edilen bir yönde bir araya getirir. Yeni deneyler, bu koşullar altında DNA uçlarının neredeyse her zaman “silme” (deletional) biçimde yeniden birleştirildiğini—aradaki bölgenin kesilip çıkarıldığını ve geride üretken, sınıf değiştirmiş bir antikor bırakıldığını—gösterdi. Ancak araştırmacılar bazı segmentleri ters çevirip karşı yöne baktırdıklarında veya başlangıç bölgesine daha yakın taşıdıklarında denge değişti. Daha kısa mesafeler ve zıt yönler, uzun DNA halkalarının hakimiyetini zayıflattı ve bunun yerine uçlar arasında daha rastgele, yerel karşılaşmaları destekledi. Bu “difüzyon” hakim durum daha fazla “tersine çevirme” (inversional) birleşim üretti; burada aradaki DNA çıkarılmak yerine ters çevrilir ve genel olarak sınıf değiştirme daha az verimli hale gelir.
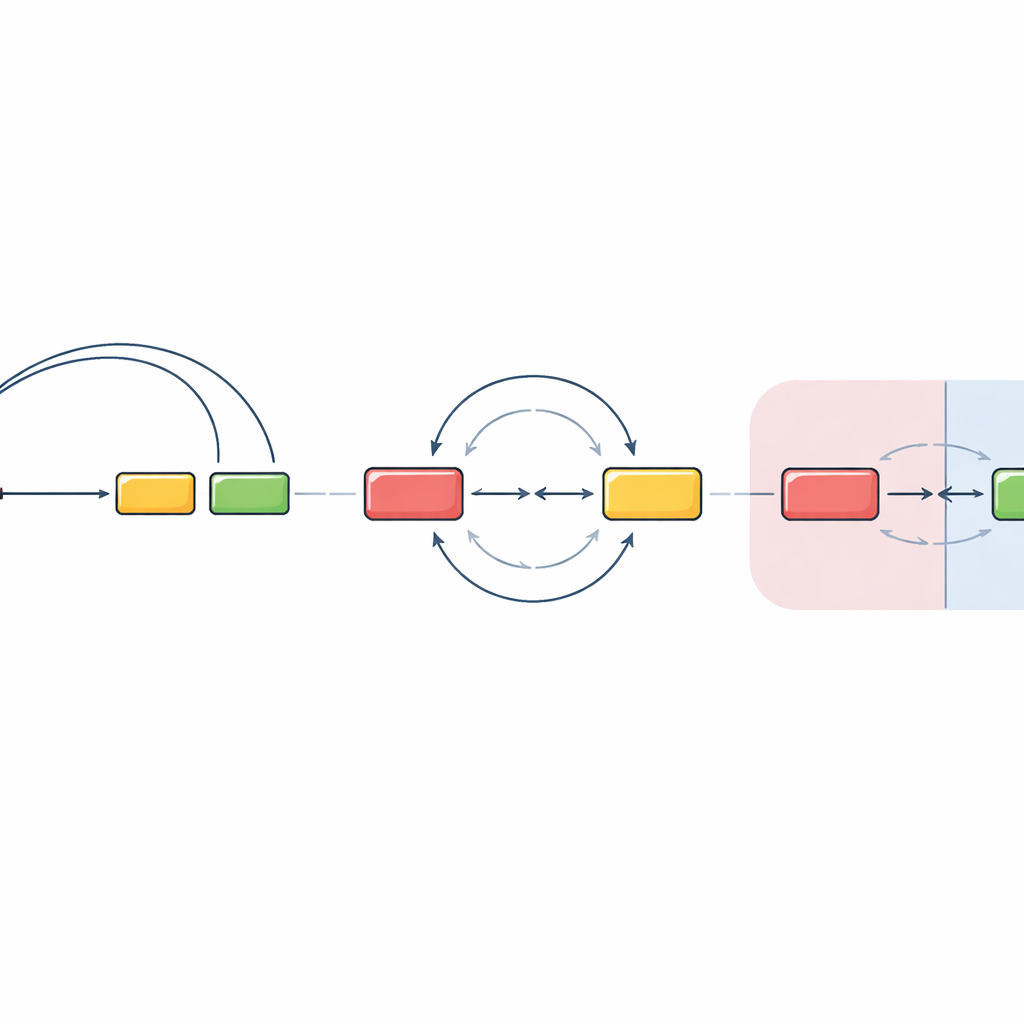
Mahalle Sınırlarını Aşmak Kuralları Değiştirir
Ekip daha sonra başlangıç ve hedef segmentlerin farklı DNA mahallelerine, yani kromatin bölgelerine yerleştirildiğinde ne olduğunu test etti. Bu bölgeler çekirdek içinde kısmen izole edilmiş ayrı odalar gibi davranır. Bir düzenleyici bölgeyi ve onun yakınındaki antikor segmentini orijinal bölgenin dışına taşıyarak, alışılmış yönlendirilmiş halkalamanın büyük ölçüde kaybolduğunu buldular. Bunun yerine, farklı bölgelerdeki DNA kırıkları çok daha yönelimsiz bir şekilde karşılaşıp yeniden birleşti; silme ve tersine çevirme olasılıkları kabaca eşitti. İlginç biçimde, ana hedef segment çıkarıldığında aynı düzenleyici mekanizmanın yakınlardaki bir “hedef dışı” DNA yamasını harekete geçirebildiğini ve bunun da benzer şekilde yönelimsiz rekombinasyona girdiğini gözlemlediler. Bu, segmentler ayrı bölgelere ayrıldığında hücrenin sıkı yönlendirilmiş halkalamaya çok fazla güvenmek yerine büyük ölçüde rastlantısal karşılaşmalara dayandığını gösterir.
Bu Gizli Mimari Neden Önemli
Toplamda, çalışma antikor genlerinin üç boyutlu düzeni ve okuma yönünün sınıf değiştirme sırasında DNA kırıklarının nasıl onarıldığını güçlü şekilde etkilediğini gösteriyor. Segmentler aynı yönü paylaşıyor, birbirlerinden uzak ve aynı bölgede yer alıyorsa, uzun DNA halkaları onları üretken birleşimlere yönlendirerek yeni antikor sınıflarını verimli biçimde üretir. Segmentler yakınsa, zıt yönlüysa veya bölgeler arasında bölünmüşse, yerel hareket ve rastlantısal karşılaşmalar devreye girer; bu da daha fazla ters çevrilmiş veya karışık sonuçlara yol açar ve genellikle daha az verimli sınıf değiştirme ile sonuçlanır. Gayri uzman bir okuyucu için ana mesaj şudur: önemli olan sadece genetik kod değil; bu kodun nasıl katlandığı, yönlendirildiği ve uzayda bölümlere ayrıldığıdır—bağışıklık sistemimizin evrimleşen patojenlerle başa çıkmak için kendi DNA’sını güvenli ve etkili biçimde yeniden yapılandırmasına yardımcı olan bir mimari mantık.
Atıf: Luo, S., Qiao, R., Zha, H. et al. Chromatin-intrinsic mechanisms determine orientation-specific class switch recombination. Nat Commun 17, 3319 (2026). https://doi.org/10.1038/s41467-026-70031-z
Anahtar kelimeler: antikor sınıf değiştirme, kromatin mimarisi, DNA halka ekstrüzyonu, B hücresi bağışıklığı, genom topolojisi