Clear Sky Science · nl
Chromatineigen mechanismen bepalen oriëntatie-specifieke klassewisselrecombinatie
Hoe ons lichaam antilichamen fijn afstemt
Wanneer we een infectie bestrijden, vertrouwt ons immuunsysteem niet op één soort antilichaam. Het kan onderdelen van antilichaammoleculen uitwisselen om beter te passen bij verschillende bedreigingen, van virussen in de longen tot bacteriën in de darmen. Dit artikel onderzoekt hoe de fysieke vouwing van DNA in immuuncellen dat uitwisselingsproces stilletjes stuurt, en zo bijdraagt aan efficiënte, nauwkeurige en passende veranderingen van antilichamen voor de behoeften van het lichaam.
Het schakelpaneel binnen antilichaamgenen
Antilichamen zijn opgebouwd uit gensegmenten die herschikt en aangepast kunnen worden als modulaire onderdelen. Vroeg in het leven van een B-cel creëert één reeks veranderingen de “kop” van het antilichaam die ziekteverwekkers herkent. Later zorgt een tweede proces, klassewisseling genoemd, voor de uitwisseling van het “staart”gebied om te veranderen hoe het antilichaam zich in het lichaam gedraagt—of het in het bloed circuleert, het darmslijmvlies bekleedt of door slijmvliesbarrières passeert. Deze wissel vindt plaats in het gen voor de zware keten van het antilichaam, waarbij een standaardsegment wordt vervangen door één van meerdere alternatieven. Hoewel al lang bekend is dat een speciaal enzym het DNA knipt om deze wissel mogelijk te maken, bleef onduidelijk waarom die knippen meestal in één richting weer met elkaar verbonden worden—en dus een bruikbaar antilichaam opleveren in plaats van een kapot of stil product.
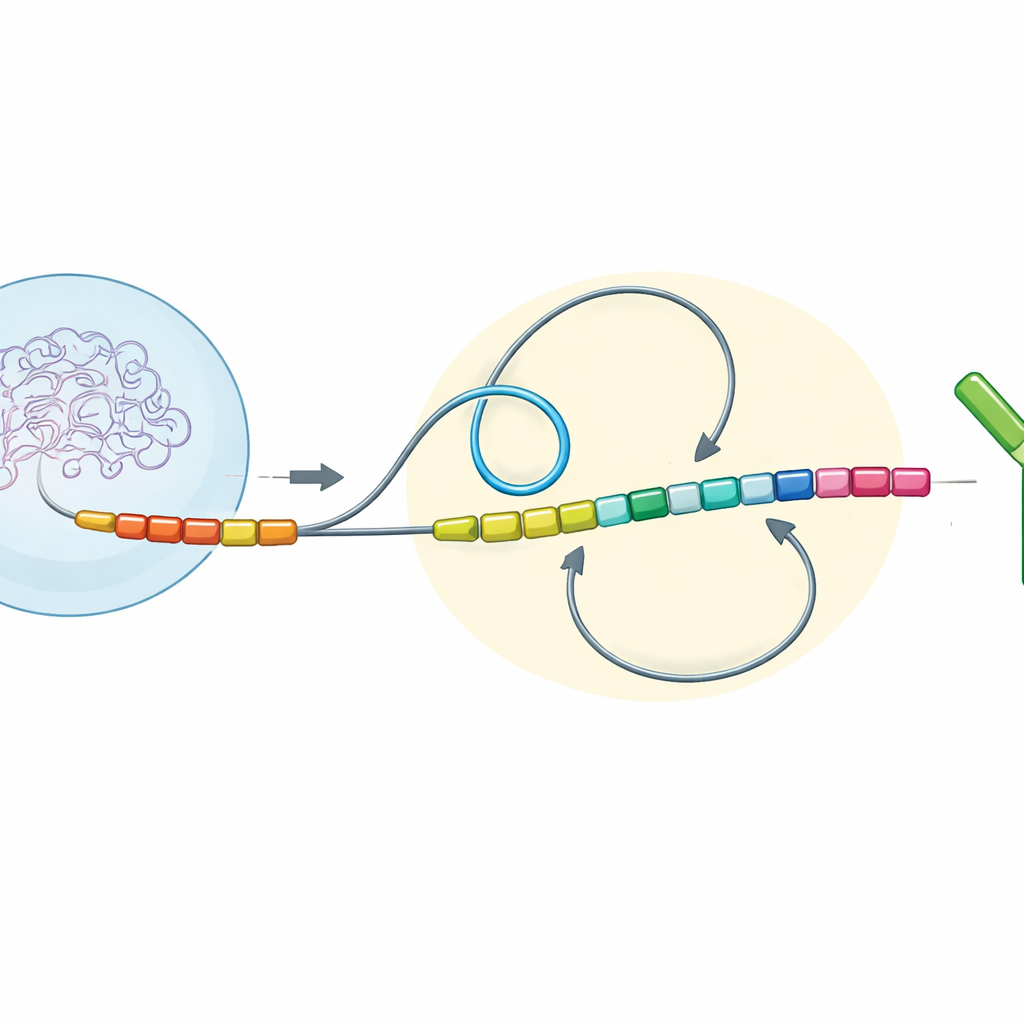
DNA-lussen en richting bepalen de uitkomst
De auteurs onderzochten antilichaamgenregio’s in veel gewervelde soorten en bouwden vervolgens gewijzigde versies van deze regio’s na in muisimmuuncellen die in het laboratorium werden gekweekt. Ze concentreerden zich op drie eenvoudige maar krachtige kenmerken van het DNA: de richting waarin verschillende segmenten worden afgelezen (transcriptieoriëntatie), hoe ver ze uit elkaar liggen langs het chromosoom (chromatine-afstand), en of ze binnen hetzelfde driedimensionale buurgebied liggen (een chromatinedomein) of in aparte gebieden. Samen noemen zij deze combinatie de “switch-topologische configuratie.” Door specifieke constantregionsegmenten nauwkeurig om te keren, te verplaatsen of te fuseren, konden ze waarnemen hoe deze wijzigingen het opnieuw verbinden van gebroken DNA-einden tijdens klassewisseling beïnvloedden.
Wanneer lange lussen de baas zijn versus wanneer lokaal geschud wint
Bij zoogdieren zoals muizen en mensen zijn de meeste antilichaam-"staart"segmenten in dezelfde richting geordend als het startsegment en relatief ver uit elkaar geplaatst langs het gen. In deze opstelling rolt een proteïnecomplex het DNA in grote lussen, waardoor de verre stukken in een bevoordeelde richting bij elkaar komen. De nieuwe experimenten toonden aan dat onder deze omstandigheden DNA-einden vrijwel altijd op een "deletionele" manier weer samengaan—het tussenliggende stuk wordt weggesneden en er blijft een productief, gewisseld antilichaam over. Maar wanneer de onderzoekers sommige segmenten omkeerden zodat ze de tegenovergestelde richting in wijzen, of ze dichterbij het startpunt plaatsten, veranderde de balans. Kortere afstanden en tegengestelde oriëntaties verzwakten de dominantie van lange DNA-lussen en gaven in plaats daarvan de voorkeur aan meer willekeurige, lokale ontmoetingen tussen einden. Deze door "diffusie" gedomineerde situatie leverde veel meer "inversionele" verbindingen op, waarbij het tussenliggende DNA wordt omgedraaid in plaats van verwijderd, en de wissel werd over het algemeen minder efficiënt.
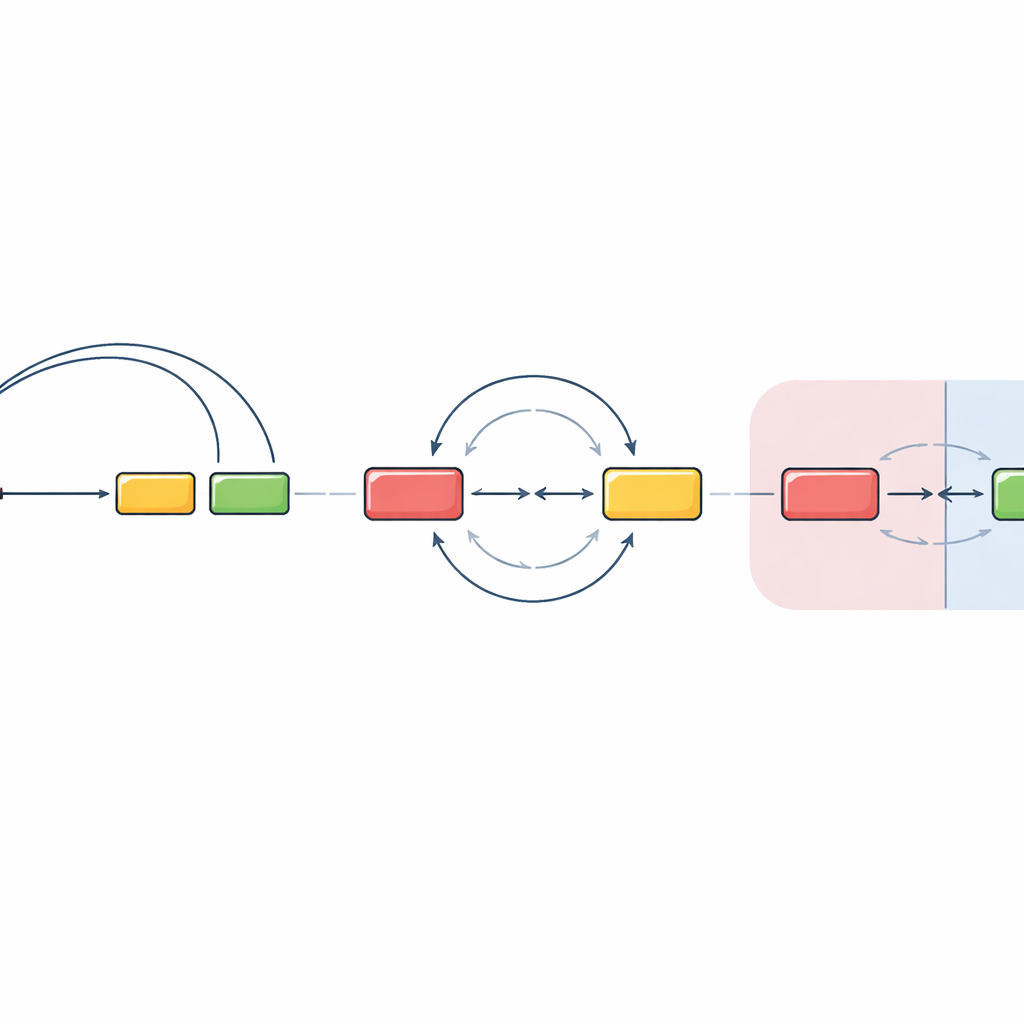
Over buurgrenzen heen verandert de spelregels
Het team testte vervolgens wat er gebeurt wanneer het start- en het doelsegment in verschillende DNA-buurten worden geplaatst, bekend als chromatinedomeinen. Deze domeinen functioneren als aparte kamers in de kern, deels geïsoleerd van elkaar. Door een belangrijke regulerende regio en het nabijgelegen antilichaamsegment buiten het oorspronkelijke domein te plaatsen, ontdekten ze dat de gebruikelijke geleide lussvorming grotendeels verdween. In plaats daarvan kwamen DNA-breuken in verschillende domeinen elkaar op een veel minder oriëntatie-gevoelige manier tegen en werden ze herverbonden, met ongeveer gelijke kansen op deletie of inversie. Interessant genoeg zagen ze ook dat wanneer het belangrijkste doelsegment werd verwijderd, dezelfde regulerende machinerie een nabijgelegen "off-target" DNA-stuk kon activeren, dat vervolgens op vergelijkbare, onvooroordeelde wijze recombinatie aanging. Dit suggereert dat zodra segmenten in afzonderlijke domeinen zitten, de cel sterk afhankelijk is van toevallige ontmoetingen in plaats van strak gerichte lussen.
Waarom deze verborgen architectuur ertoe doet
Alles bij elkaar toont het werk aan dat de driedimensionale lay-out en de lezingrichting van antilichaamgenen sterk beïnvloeden hoe DNA-breuken worden gerepareerd tijdens klassewisseling. Wanneer segmenten dezelfde richting delen, ver uit elkaar liggen en in hetzelfde domein zitten, sturen lange DNA-lussen ze naar productieve verbindingen die efficiënt nieuwe antilichaamklassen genereren. Wanneer segmenten dichtbij, tegengesteld georiënteerd of verdeeld over domeinen zijn, nemen lokale beweging en toevallige ontmoetingen het over, wat leidt tot meer omgedraaide of gemengde uitkomsten en vaak minder efficiënte wisseling. Voor de niet-specialistische lezer is de kernboodschap dat niet alleen de genetische code van belang is, maar ook hoe die code wordt gevouwen, georiënteerd en gecompartimenteerd in de ruimte—een architectonische logica die ons immuunsysteem helpt zijn eigen DNA veilig en effectief te herschakelen om gelijke tred te houden met evoluerende pathogenen.
Bronvermelding: Luo, S., Qiao, R., Zha, H. et al. Chromatin-intrinsic mechanisms determine orientation-specific class switch recombination. Nat Commun 17, 3319 (2026). https://doi.org/10.1038/s41467-026-70031-z
Trefwoorden: antilichaamklassewisseling, chromatine-architectuur, DNA-lusextrusie, B-celimmuniteit, genoomtopologie