Clear Sky Science · en
Chromatin-intrinsic mechanisms determine orientation-specific class switch recombination
How Our Bodies Fine-Tune Antibodies
When we fight infections, our immune system doesn’t rely on just one kind of antibody. It can swap out parts of antibody molecules to better match different threats, from viruses in the lungs to bacteria in the gut. This paper explores how the physical folding of DNA inside immune cells quietly steers that swapping process, helping ensure that antibody changes are efficient, accurate, and suited to the body’s needs.
The Switchboard Inside Antibody Genes
Antibodies are built from gene segments that can be rearranged and refitted like modular pieces. Early in a B cell’s life, one set of changes creates the “head” of the antibody that recognizes germs. Later, a second process, called class switching, swaps the “tail” region to change how the antibody behaves in the body—whether it circulates in blood, lines the gut, or crosses mucosal surfaces. This swap occurs in the antibody heavy-chain gene, where a default segment is replaced by one of several alternatives. While it has long been known that a dedicated enzyme cuts the DNA to allow this swap, why those cuts usually reconnect in one orientation—and thus give a useful antibody rather than a broken or silent product—has remained puzzling.
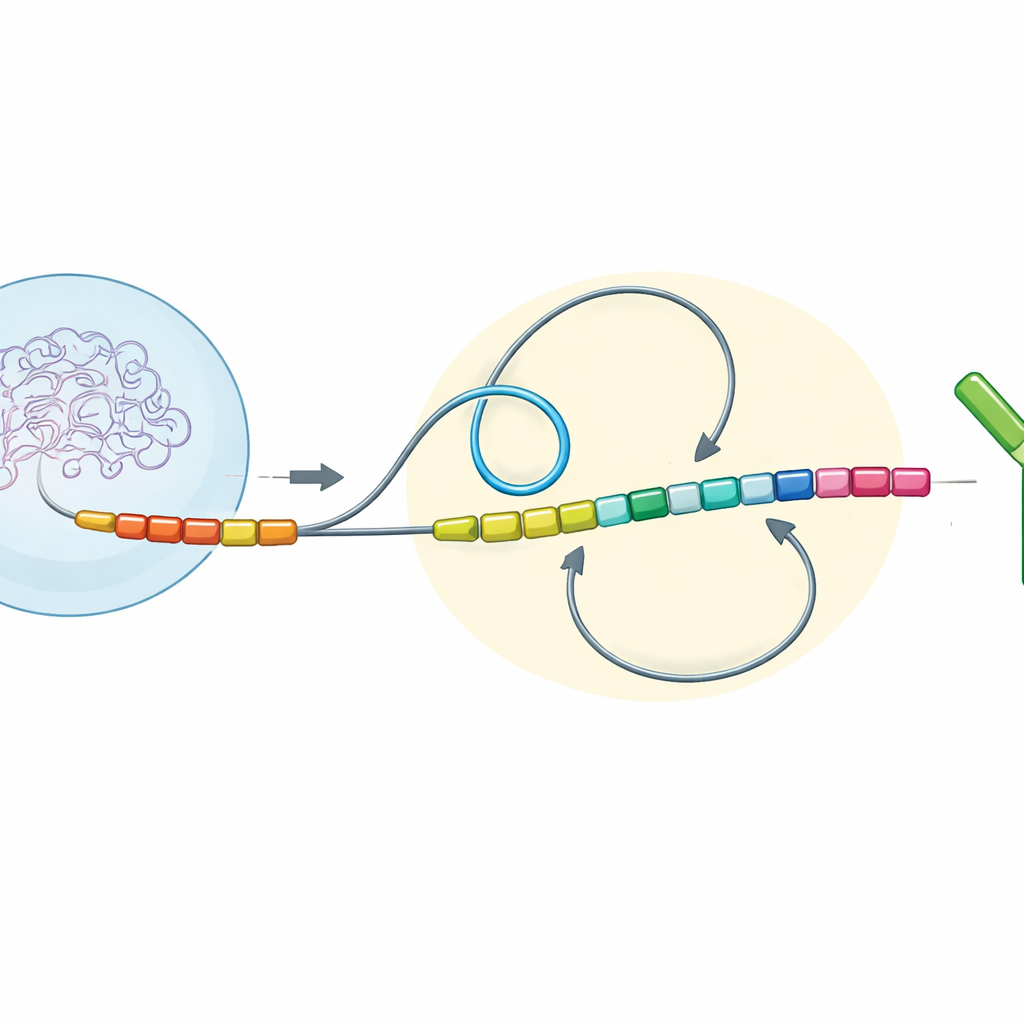
DNA Loops and Direction Shape the Outcome
The authors examined antibody gene regions across many vertebrate species and then rebuilt altered versions of these regions in mouse immune cells grown in the lab. They focused on three simple but powerful features of the DNA: the direction in which different segments are read (transcription orientation), how far apart they sit along the chromosome (chromatin distance), and whether they lie inside the same three-dimensional neighborhood (a chromatin domain) or in separate ones. Together, they call this combination the “switch topological configuration.” By precisely flipping, moving, or fusing specific constant-region segments, they could watch how these changes affected the way broken DNA ends rejoined during class switching.
When Long Loops Rule vs. When Local Jostling Wins
In mammals such as mice and humans, most antibody “tail” segments are arranged facing the same way as the starting segment and are spaced relatively far apart along the gene. In this setup, a protein complex reels the DNA into large loops, bringing the distant pieces together in a favored direction. The new experiments showed that, under these conditions, DNA ends nearly always rejoin in a “deletional” way—cutting out the intervening stretch and leaving behind a productive, switched antibody. But when the researchers flipped some segments so they faced the opposite direction, or moved them closer to the start site, the balance changed. Shorter distances and opposing directions weakened the dominance of long DNA loops and instead favored more random, local encounters between ends. This “diffusion”-dominated situation produced many more “inversional” joins, where the intervening DNA is flipped rather than removed, and switching became less efficient overall.
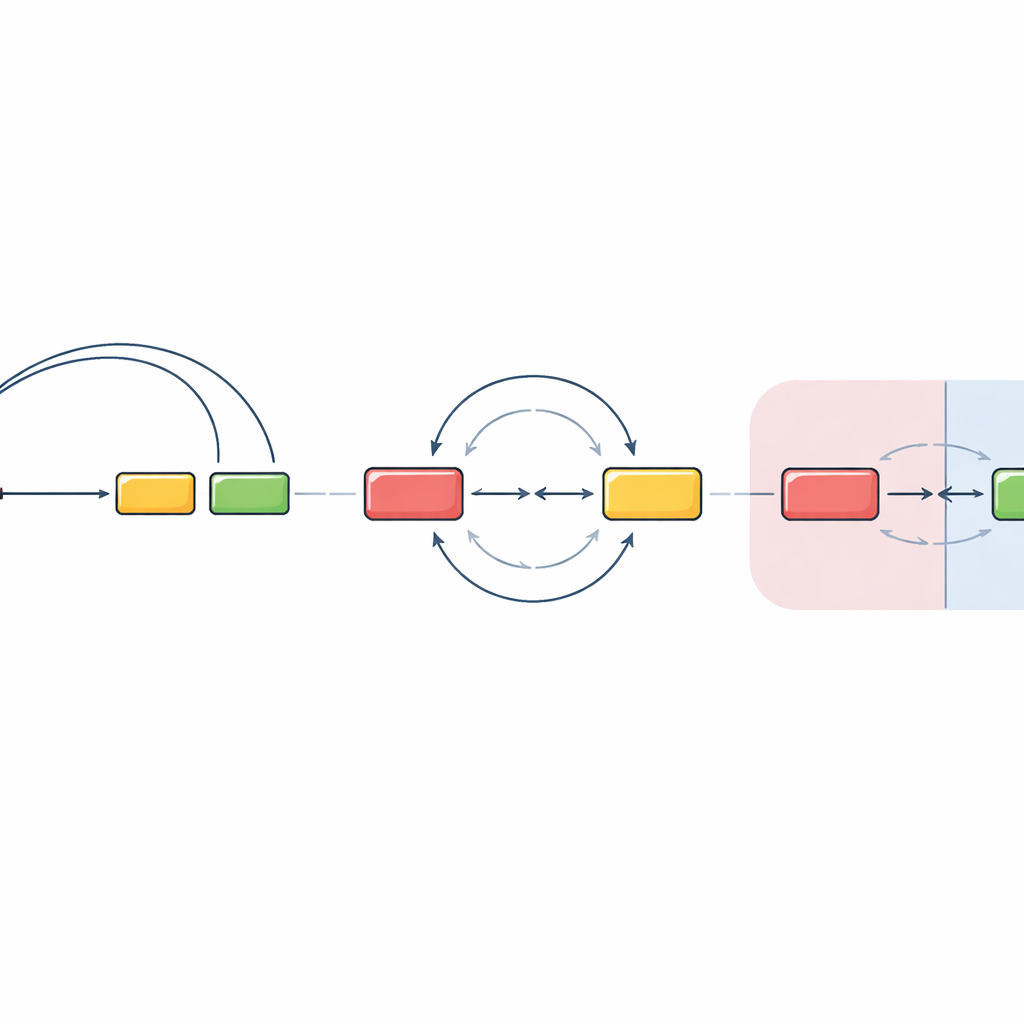
Crossing Neighborhood Lines Changes the Rules
The team then tested what happens when the starting and target segments are placed in different DNA neighborhoods, known as chromatin domains. These domains act like separate rooms inside the nucleus, partly insulated from each other. By relocating a key regulatory region and its nearby antibody segment outside the original domain, they found that the usual guided looping was largely lost. Instead, DNA breaks in different domains met and rejoined in a much more orientation-unbiased way, with roughly equal chances of deletion or inversion. Interestingly, they also observed that when the main target segment was removed, the same regulatory machinery could awaken a nearby “off-target” DNA patch, which then entered into similarly unbiased recombination. This suggests that once segments are separated into distinct domains, the cell relies heavily on chance encounters rather than tightly directed looping.
Why This Hidden Architecture Matters
Altogether, the work shows that the three-dimensional layout and reading direction of antibody genes strongly influence how DNA breaks are repaired during class switching. When segments share the same direction, are far apart, and sit in the same domain, long DNA loops guide them into productive joins that efficiently generate new antibody classes. When segments are close, oppositely oriented, or split between domains, local motion and chance encounters take over, leading to more flipped or mixed outcomes and often less efficient switching. For a lay reader, the key message is that it’s not just the genetic code that matters, but also how that code is folded, oriented, and compartmentalized in space—an architectural logic that helps our immune system safely and effectively rewire its own DNA to keep pace with evolving pathogens.
Citation: Luo, S., Qiao, R., Zha, H. et al. Chromatin-intrinsic mechanisms determine orientation-specific class switch recombination. Nat Commun 17, 3319 (2026). https://doi.org/10.1038/s41467-026-70031-z
Keywords: antibody class switching, chromatin architecture, DNA loop extrusion, B cell immunity, genome topology