Clear Sky Science · ru
Внутренние хроматиновые механизмы определяют ориентационно-специфичную перестройку класса антител
Как наши тела точно настраивают антитела
Когда мы боремся с заражениями, наша иммунная система не полагается на один вид антител. Она может заменять части молекул антител, чтобы лучше соответствовать разным угрозам — от вирусов в лёгких до бактерий в кишечнике. В этой статье исследуется, как физическое свёртывание ДНК внутри иммунных клеток тихо направляет этот процесс замены, помогая обеспечивать эффективность, точность и соответствие изменений антител потребностям организма.
Переключатель внутри генов антител
Антитела состоят из генетических сегментов, которые можно переставлять и подгонять как модульные детали. Рано в жизни В-клетки один набор перестроек формирует «головку» антитела, которая распознаёт чужеродные молекулы. Позже второй процесс, называемый переключением класса, меняет «хвостовую» область, чтобы изменить поведение антитела в организме — будет ли оно циркулировать в крови, выстилать кишечник или пересекать слизистые поверхности. Эта замена происходит в гене тяжёлой цепи антител, где сегмент по умолчанию заменяется одним из нескольких альтернативных. Хотя давно известно, что специализированный фермент разрезает ДНК, чтобы позволить эту замену, сохраняется загадка: почему эти разрывы обычно вновь соединяются в одном направлении — и таким образом дают полезное антитело, а не разорванный или немой продукт.
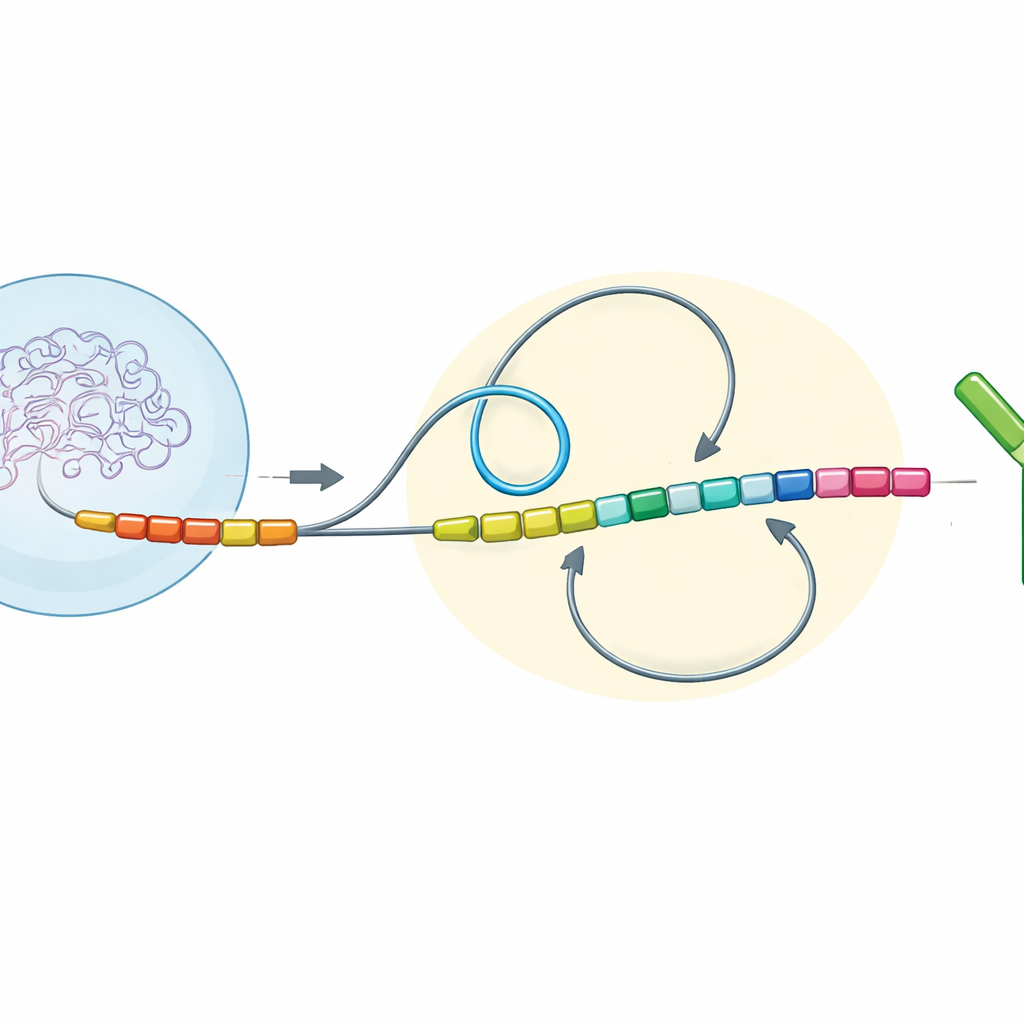
Петли ДНК и направление задают результат
Авторы изучили регионы генов антител у многих позвоночных и затем восстановили изменённые версии этих участков в иммунных клетках мыши, выращенных в лаборатории. Они сосредоточились на трёх простых, но мощных признаках ДНК: направлении чтения разных сегментов (ориентация транскрипции), расстоянии между ними вдоль хромосомы (хроматиновое расстояние) и том, находятся ли они внутри одного трёхмерного соседства (хроматинового домена) или в разных. Вместе они называют эту комбинацию «топологической конфигурацией переключения». Точно переворачивая, перемещая или сливая конкретные сегменты константной области, они могли наблюдать, как эти изменения влияют на то, как разорванные концы ДНК вновь соединяются во время переключения класса.
Когда правят длинные петли и когда побеждает локальная суетливость
У млекопитающих, таких как мыши и люди, большинство «хвостовых» сегментов антител ориентированы так же, как исходный сегмент, и расположены относительно далеко друг от друга вдоль гена. В такой конфигурации белковый комплекс сматывает ДНК в большие петли, сводя отдалённые участки вместе в предпочтительном направлении. Новые эксперименты показали, что в этих условиях концы ДНК почти всегда вновь соединяются «делеционно» — промежуточный участок вырезается, и остаётся продуктивное, переключённое антитело. Но когда исследователи переворачивали некоторые сегменты так, что они были направлены в противоположную сторону, или приближали их к стартовой точке, баланс менялся. Короткие расстояния и противоположная ориентация ослабляли доминирование длинных ДНК-петель и вместо этого благоволили более случайным локальным встречам между концами. Эта «диффузионно»-доминированная ситуация приводила к гораздо большему числу «инверсионных» соединений, где промежуточная ДНК переворачивается, а не удаляется, и переключение в целом становилось менее эффективным.
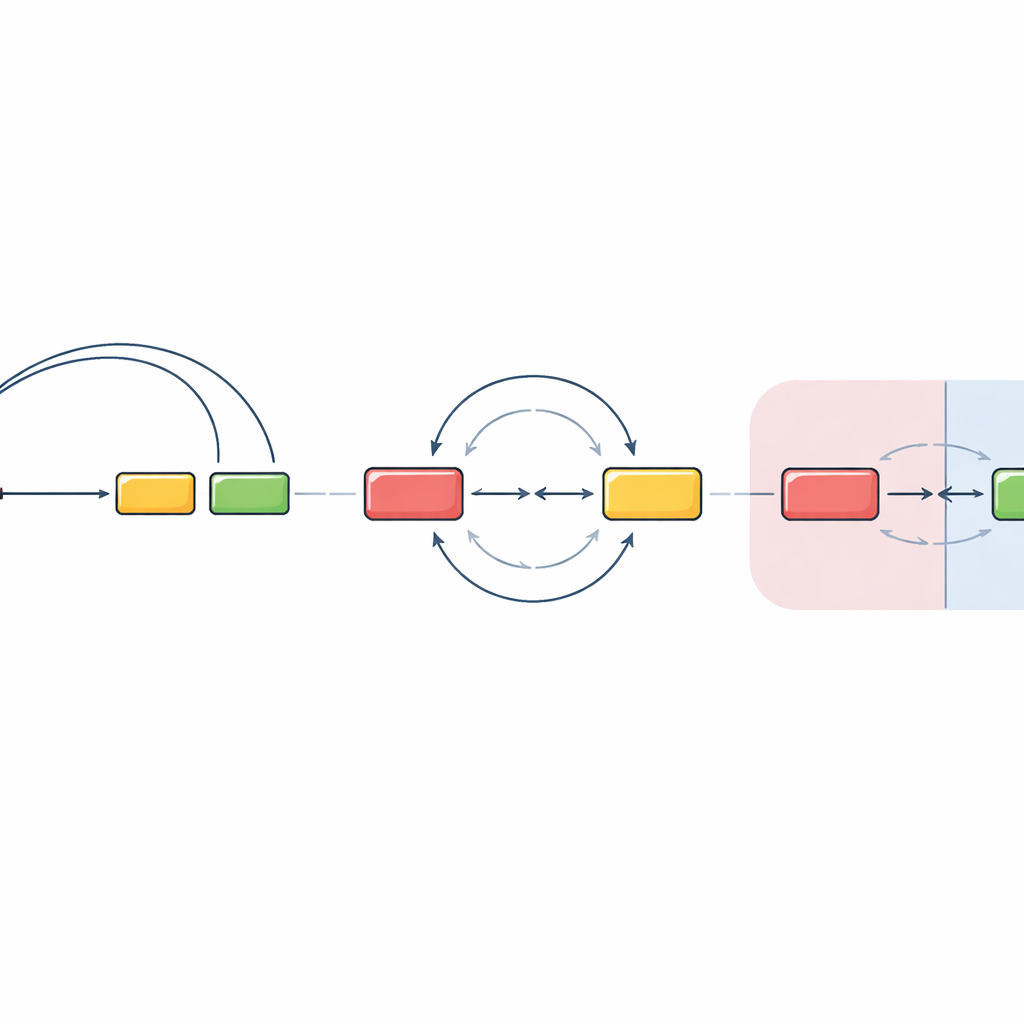
Пересечение границ соседств меняет правила
Команда затем проверила, что происходит, когда исходный и целевой сегменты помещают в разные ДНК-соседства, известные как хроматиновые домены. Эти домены действуют как отдельные комнаты внутри ядра, частично изолированные друг от друга. Переместив ключевую регуляторную область и прилежащий сегмент антитела за пределы исходного домена, они обнаружили, что обычное направляемое образование петель в значительной степени теряется. Вместо этого разрывы ДНК в разных доменах встречались и вновь соединялись гораздо более свободно по ориентации, с приблизительно равными шансами на делекцию или инверсию. Интересно, что они также заметили: когда главный целевой сегмент удаляли, та же регуляторная машина могла «пробудить» близлежащий «внецелевой» участок ДНК, который затем вступал в аналогично бесспреферентную рекомбинацию. Это указывает на то, что как только сегменты оказываются в разных доменах, клетка в значительной мере полагается на случайные встречи, а не на строго направляемые петли.
Почему эта скрытая архитектура важна
В совокупности работа показывает, что трёхмерная компоновка и направление чтения генов антител сильно влияют на то, как ремонтируются разрывы ДНК во время переключения класса. Когда сегменты имеют одинаковую ориентацию, находятся далеко друг от друга и расположены в одном домене, длинные ДНК-петли направляют их в продуктивные соединения, которые эффективно генерируют новые классы антител. Когда сегменты близки, противоположно ориентированы или разделены между доменами, локальное движение и случайные встречи берут верх, приводя к большему числу перевёрнутых или смешанных исходов и часто к менее эффективному переключению. Для неспециалиста ключевая мысль в том, что важно не только генетический код, но и то, как этот код сложен, ориентирован и разделён в пространстве — архитектурная логика, которая помогает нашей иммунной системе безопасно и эффективно перестраивать собственную ДНК, чтобы поспевать за эволюционирующими патогенами.
Цитирование: Luo, S., Qiao, R., Zha, H. et al. Chromatin-intrinsic mechanisms determine orientation-specific class switch recombination. Nat Commun 17, 3319 (2026). https://doi.org/10.1038/s41467-026-70031-z
Ключевые слова: переключение класса антител, архитектура хроматина, выворачивание ДНК в петли, иммунитет B-клеток, топология генома